Структура одного білка визначається групою генів. Білки: первинна структура білків, схема утворення трипептиду
Білки- Високомолекулярні органічні сполуки, що складаються із залишків α-амінокислот.
У склад білківвходять вуглець, водень, азот, кисень, сірка. Частина білків утворює комплекси з іншими молекулами, які містять фосфор, залізо, цинк та мідь.
Білки мають велику молекулярну масу: яєчний альбумін — 36 000, гемоглобін — 152 000, міозин — 500 000. Для порівняння: молекулярна маса спирту — 46, оцтової кислоти — 60, бензолу — 78.
Амінокислотний склад білків
Білки- Неперіодичні полімери, мономерами яких є α-амінокислоти. Зазвичай як мономери білків називають 20 видів α-амінокислот, хоча в клітинах і тканинах їх виявлено понад 170.
Залежно від того, чи можуть амінокислоти синтезуватися в організмі людини та інших тварин, розрізняють: замінні амінокислоти- Можуть синтезуватися; незамінні амінокислоти- Не можуть синтезуватися. Незамінні амінокислоти повинні надходити в організм разом із їжею. Рослини синтезують усі види амінокислот.
Залежно від амінокислотного складу, білки бувають: повноцінними- Містять весь набір амінокислот; неповноцінними- Якісь амінокислоти в їхньому складі відсутні. Якщо білки складаються лише з амінокислот, їх називають простими. Якщо білки містять окрім амінокислот ще й неамінокислотний компонент (простетичну групу), їх називають складними. Простетична група може бути представлена металами (металопротеїни), вуглеводами (глікопротеїни), ліпідами (ліпопротеїни), нуклеїновими кислотами (нуклеопротеїни).
Усе амінокислоти містять: 1) карбоксильну групу (-СООН); 2) аміногрупу (-NH 2); 3) радикал або R-групу (решта молекули). Будова радикала у різних видів амінокислот різна. Залежно від кількості аміногруп та карбоксильних груп, що входять до складу амінокислот, розрізняють: нейтральні амінокислоти, Що мають одну карбоксильну групу та одну аміногрупу; основні амінокислоти, Що мають більше однієї аміногрупи; кислі амінокислоти, Що мають більше однієї карбоксильної групи
Амінокислоти є амфотерними сполуками, Так як у розчині вони можуть виступати як у ролі кислот, так і основ. У водних розчинах амінокислоти існують у різних іонних формах.
Пептидна зв'язок
Пептиди- Органічні речовини, що складаються з залишків амінокислот, з'єднаних пептидним зв'язком.
Утворення пептидів відбувається внаслідок реакції конденсації амінокислот. При взаємодії аміногрупи однієї амінокислоти з карбоксильною групою іншою між ними виникає ковалентний азот-вуглецевий зв'язок, яку і називають пептидний. Залежно кількості амінокислотних залишків, що входять до складу пептиду, розрізняють дипептиди, трипептиди, тетрапептидиі т.д. Утворення пептидного зв'язку може повторюватися багаторазово. Це призводить до освіти поліпептидів. На одному кінці пептиду знаходиться вільна аміногрупа (його називають N-кінцем), а на іншому - вільна карбоксильна група (його називають С-кінцем).

Просторова організація білкових молекул
Виконання білками певних специфічних функцій залежить від просторової конфігурації їх молекул, крім того, клітині енергетично невигідно тримати білки у розгорнутій формі, у вигляді ланцюжка, тому поліпептидні ланцюги піддаються укладання, набуваючи певної тривимірної структури, або конформації. Виділяють 4 рівні просторової організації білків.
Первинна структура білка- Послідовність розташування амінокислотних залишків у поліпептидному ланцюгу, що становить молекулу білка. Зв'язок між амінокислотами - пептидна.

Якщо молекула білка складається всього з 10 амінокислотних залишків, то теоретично можливих варіантів білкових молекул, що відрізняються порядком чергування амінокислот, — 10 20 . Маючи 20 амінокислот, можна скласти з них ще більшу кількість різноманітних комбінацій. В людини виявлено близько десяти тисяч різних білків, які відрізняються як один від одного, так і від білків інших організмів.
Саме первинна структура білкової молекули визначає властивості молекул білка та її просторову конфігурацію. Заміна лише однієї амінокислоти на іншу в поліпептидному ланцюжку призводить до зміни властивостей і функцій білка. Наприклад, заміна в β-субодиниці гемоглобіну шостої глутамінової амінокислоти на валін призводить до того, що молекула гемоглобіну в цілому не може виконувати свою основну функцію - транспорт кисню; у таких випадках у людини розвивається захворювання – серповидноклітинна анемія.
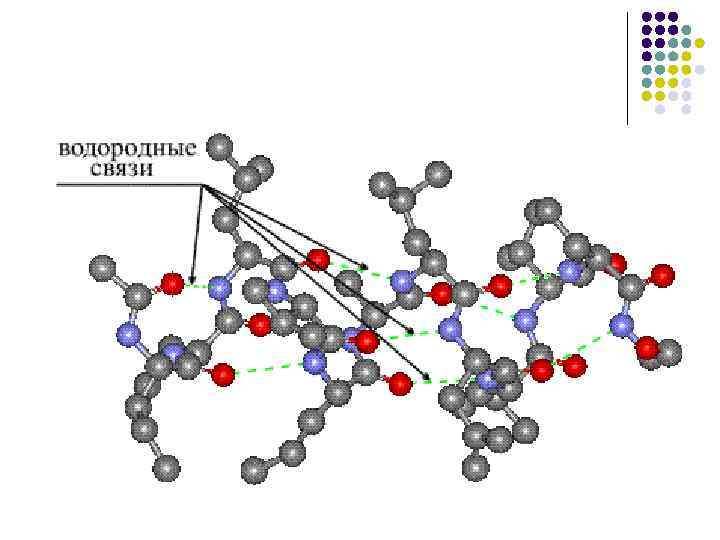
Вторинна структура- Упорядковане згортання поліпептидного ланцюга в спіраль (має вигляд розтягнутої пружини). Витки спіралі зміцнюються водневими зв'язками, що виникають між карбоксильними групами та аміногрупами. Практично всі СО- та NН-групи беруть участь в утворенні водневих зв'язків. Вони слабші пептидних, але, повторюючись багаторазово, надають цієї зміни стійкість і жорсткість. На рівні вторинної структури існують білки: фіброїн (шовк, павутиння), кератин (волосся, нігті), колаген (сухожилля).



Третинна структура- укладання поліпептидних ланцюгів у глобули, що виникає внаслідок виникнення хімічних зв'язків (водневих, іонних, дисульфідних) та встановлення гідрофобних взаємодій між радикалами амінокислотних залишків. Основну роль освіті третинної структури грають гидрофильно-гидрофобные взаємодії. У водних розчинах гідрофобні радикали прагнуть сховатися від води, групуючись усередині глобули, тоді як гідрофільні радикали в результаті гідратації (взаємодії з диполями води) прагнуть опинитися на поверхні молекули. У деяких білків третинна структура стабілізується дисульфідними ковалентними зв'язками, що виникають між атомами сірки двох залишків цистеїну. На рівні третинної структури є ферменти, антитіла, деякі гормони.

Четвертична структурай у складних білків, молекули яких утворені двома і більше глобулами. Субодиниці утримуються в молекулі завдяки іонним, гідрофобним та електростатичним взаємодіям. Іноді при утворенні четвертинної структури між субодиницями виникають дисульфідні зв'язки. Найбільш вивченим білком, що має четвертинну структуру, є гемоглобін. Він утворений двома α-субодиницями (141 амінокислотний залишок) і двома β-субодиницями (146 амінокислотних залишків). З кожною субодиницею пов'язана молекула гема, що містить залізо.
Якщо з будь-яких причин просторова конформація білків відхиляється від нормальної, білок не може виконувати своїх функцій. Наприклад, причиною «коров'ячого сказу» (губкоподібної енцефалопатії) є аномальна конформація пріонів – поверхневих білків нервових клітин.
Властивості білків
Амінокислотний склад, структура білкової молекули визначають його властивості. Білки поєднують у собі основні та кислотні властивості, що визначаються радикалами амінокислот: чим більше кислих амінокислот у білку, тим яскравіше виражені його кислотні властивості. Здатність віддавати та приєднувати Н+ визначають буферні властивості білків; один із найпотужніших буферів - гемоглобін в еритроцитах, що підтримує рН крові на постійному рівні. Є розчинні білки (фібриноген), є нерозчинні, що виконують механічні функції (фіброїн, кератин, колаген). Є активні білки в хімічному відношенні (ферменти), є хімічно неактивні, стійкі до впливу різних умов зовнішнього середовища і вкрай нестійкі.
Зовнішні фактори (нагрівання, ультрафіолетове випромінювання, важкі метали та їх солі, зміни рН, радіація, зневоднення)
можуть спричиняти порушення структурної організації молекули білка. Процес втрати тривимірної конформації, властивої даної молекули білка, називають денатурацією. Причиною денатурації є розрив зв'язків, що стабілізують певну структуру білка. Спочатку рвуться найслабші зв'язки, а при посиленні умов і сильніші. Тому спочатку втрачається четвертинна, потім третинна та вторинна структури. Зміна просторової конфігурації призводить до зміни властивостей білка і, як наслідок, унеможливлює виконання білком властивих йому біологічних функцій. Якщо денатурація не супроводжується руйнуванням первинної структури, вона може бути оборотний, у разі відбувається самовідновлення властивої білку конформації. Такої денатурації зазнають, наприклад, рецепторні білки мембрани. Процес відновлення структури білка після денатурації називається ренатурацією. Якщо відновлення просторової конфігурації білка неможливе, то денатурація називається незворотній.
Функції білків
| Функція | Приклади та пояснення |
|---|---|
| Будівельна | Білки беруть участь в утворенні клітинних та позаклітинних структур: входять до складу клітинних мембран (ліпопротеїни, глікопротеїни), волосся (кератин), сухожиль (колаген) тощо. |
| Транспортна | Білок крові гемоглобін приєднує кисень і транспортує його від легень до всіх тканин та органів, а від них у легені переносить вуглекислий газ; До складу клітинних мембран входять особливі білки, які забезпечують активне і строго вибіркове перенесення деяких речовин та іонів з клітини у зовнішнє середовище і назад. |
| Регуляторна | Гормони білкової природи беруть участь у регулюванні процесів обміну речовин. Наприклад, гормон інсулін регулює рівень глюкози в крові, сприяє синтезу глікогену, збільшує утворення жирів із вуглеводів. |
| Захисна | У у відповідь проникнення в організм чужорідних білків чи мікроорганізмів (антигенів) утворюються особливі білки — антитіла, здатні пов'язувати і знешкоджувати їх. Фібрин, що утворюється з фібриногену, сприяє зупинці кровотеч. |
| Двигуна | Скорочувальні білки актин та міозин забезпечують скорочення м'язів у багатоклітинних тварин. |
| Сигнальна | У поверхневу мембрану клітини вбудовані молекули білків, здатних змінювати свою третинну структуру у відповідь на дію факторів зовнішнього середовища, таким чином здійснюючи прийом сигналів із зовнішнього середовища та передачу команд у клітину. |
| Запасаюча | В організмі тварин білки, як правило, не запасаються, виняток: альбумін яєць, казеїн молока. Але завдяки білкам в організмі можуть відкладатися про запас деякі речовини, наприклад, при розпаді гемоглобіну залізо не виводиться з організму, а зберігається, утворюючи комплекс з ферритином білком. |
| Енергетична | При розпаді 1 г білка до кінцевих продуктів виділяється 176 кДж. Спочатку білки розпадаються до амінокислот, а потім до кінцевих продуктів – води, вуглекислого газу та аміаку. Однак як джерело енергії білки використовуються тільки тоді, коли інші джерела (вуглеводи та жири) витрачені. |
| Каталітична | Одна з найважливіших функцій білків. Забезпечується білками - ферментами, які прискорюють біохімічні реакції, що відбуваються у клітинах. Наприклад, рибульозобіфосфаткарбоксилаза каталізує фіксацію 2 при фотосинтезі. |
Ферменти
Ферменти, або ензими, - Особливий клас білків, що є біологічними каталізаторами. Завдяки ферментам біохімічні реакції протікають із величезною швидкістю. Швидкість ферментативних реакцій у десятки тисяч разів (а іноді й у мільйони) вища за швидкість реакцій, що йдуть за участю неорганічних каталізаторів. Речовина, на яку надає свою дію фермент, називають субстратом.
Ферменти - глобулярні білки, особливостям будовиферменти можна розділити на дві групи: прості та складні. Прості ферментиє найпростішими білками, тобто. складаються лише з амінокислот. Складні ферментиє складними білками, тобто. до їх складу, крім білкової частини, входить група небілкової природи. кофактор. У деяких ферментів як кофактор виступають вітаміни. У молекулі ферменту виділяють особливу частину, яка називається активним центром. Активний центр- Невелика ділянка ферменту (від трьох до дванадцяти амінокислотних залишків), де і відбувається зв'язування субстрату або субстратів з утворенням фермент-субстратного комплексу. Після завершення реакції фермент-субстратний комплекс розпадається на фермент та продукт (продукти) реакції. Деякі ферменти мають (крім активного) алостеричні центри- Ділянки, до яких приєднуються регулятори швидкості роботи ферменту ( алостеричні ферменти).

Для реакцій ферментативного каталізу характерні: 1) висока ефективність, 2) строга вибірковість та спрямованість дії, 3) субстратна специфічність, 4) тонка та точна регуляція. Субстратну та реакційну специфічність реакцій ферментативного каталізу пояснюють гіпотези Е. Фішера (1890 р.) та Д. Кошланда (1959 р.).
Е. Фішер (гіпотеза «ключ-замок»)припустив, що просторові конфігурації активного центру ферменту та субстрату повинні точно відповідати один одному. Субстрат порівнюється з «ключом», фермент – із «замком».
Д. Кошланд (гіпотеза «рука-рукавичка»)припустив, що просторова відповідність структури субстрату та активного центру ферменту створюється лише в момент їхньої взаємодії один з одним. Цю гіпотезу ще називають гіпотезою індукованої відповідності.
Швидкість ферментативних реакцій залежить від: 1) температури; 2) концентрації ферменту; 3) концентрації субстрату; 4) рН. Слід наголосити, що оскільки ферменти є білками, то їх активність найбільш висока за фізіологічно нормальних умов.
Більшість ферментів може працювати лише за температури від 0 до 40 °С. У цих межах швидкість реакції підвищується приблизно 2 рази при підвищенні температури на кожні 10 °С. При температурі вище 40 ° С білок піддається денатурації та активність ферменту падає. При температурі, близькій до точки замерзання, ферменти інактивуються.
При збільшенні кількості субстрату швидкість ферментативної реакції зростає до тих пір, поки кількість молекул субстрату стане рівною кількості молекул ферменту. При подальшому збільшенні кількості субстрату швидкість не збільшуватиметься, оскільки відбувається насичення активних центрів ферменту. Збільшення концентрації ферменту призводить до посилення каталітичної активності, так як в одиницю часу перетворень піддається більша кількість молекул субстрату.

Для кожного ферменту існує оптимальне значення рН, при якому він виявляє максимальну активність (пепсин – 2,0, амілаза слини – 6,8, ліпаза підшлункової залози – 9,0). При більш високих чи низьких значеннях рН активність ферменту знижується. При різких зрушеннях рН фермент денатурує.
Швидкість роботи алостеричних ферментів регулюється речовинами, що приєднуються до алостеричних центрів. Якщо ці речовини прискорюють реакцію, вони називаються активаторами, якщо гальмують інгібіторами.
Класифікація ферментів
За типом каталізованих хімічних перетворень ферменти поділені на 6 класів:
- оксиредуктази(перенесення атомів водню, кисню або електронів від однієї речовини до іншої - дегідрогеназу),
- трансферази(перенесення метильної, ацильної, фосфатної або аміногрупи від однієї речовини до іншої - трансаміназа),
- гідролази(Реакції гідролізу, при яких з субстрату утворюються два продукти - амілаза, ліпаза),
- ліази(негідролітичне приєднання до субстрату або відщеплення від нього групи атомів, при цьому можуть розриватися зв'язки С-С, С-N, С-О, С-S декарбоксилаза),
- ізомерази(Внутрішньомолекулярна перебудова - ізомераза),
- лігази(Сполука двох молекул в результаті утворення зв'язків С-С, С-N, С-О, С-S - синтетаза).
Класи у свою чергу поділені на підкласи та підпідкласи. У міжнародній класифікації, що діє, кожен фермент має певний шифр, що складається з чотирьох чисел, розділених точками. Перше число – клас, друге – підклас, третє – підпідклас, четверте – порядковий номер ферменту у цьому підпідкласі, наприклад, шифр аргінази – 3.5.3.1.
Перейти до лекції №2«Будова та функції вуглеводів та ліпідів»
Перейти до лекції №4«Будова та функції нуклеїнових кислот АТФ»
Біосинтез білка.
1. Структура одного білка визначається:
1) групою генів 2) одним геном
3) однією молекулою ДНК 4) сукупністю генів організму
2. Ген кодує інформацію про послідовність мономерів у молекулі:
1) т-РНК 2) АК 3) глікогену 4) ДНК
3. Антикодонами називаються триплети:
1)ДНК 2) т-РНК 3)і-РНК 4) р-РНК
4. Пластичний обмін складається переважно з реакцій:
1) розпаду органічних речовин 2) розпаду неорганічних речовин
3) синтез органічних речовин 4) синтез неорганічних речовин
5. Синтез білка у прокаріотичній клітині відбувається:
1) на рибосомах в ядрі 2) на рибосомах у цитоплазмі 3) у клітинній стінці
4) на зовнішній поверхні цитоплазматичної мембрани
6. Процес трансляції відбувається:
1) у цитоплазмі 2) у ядрі 3) у мітохондріях
4) на мембранах шорсткої ендоплазматичної мережі
7. На мембранах гранулярної ендоплазматичної мережі відбувається синтез:
1) АТФ; 2) вуглеводів; 3) ліпідів; 4) білків.
8. Один триплет кодує:
1. одну АК 2 одна ознака організму 3. кілька АК
9. Синтез білка завершується у момент
1. впізнавання кодону антикодоном 2. поява на рибосомі «розділового знака»
3.надходження і-РНК на рибосому
10. Процес, внаслідок якого відбувається зчитування інформації з молекули ДНК.
1.трансляція 2.транскрипція 3.трансформація
11. Властивості білків визначається…
1.вторинною структурою білка 2.первинною структурою білка
3. третинною структурою білка
12. Процес, при якому антикодон дізнається кодон на і-РНК
13. Етапи біосинтезу білка.
1.транскрипція, трансляція 2.трансформація, трансляція
3.трансорганізація, транскрипція
14. Антикодон т-РНК складається із нуклеотидів УЦГ. Який триплет ДНК йому комплементарний?
1.УУГ 2. ТТЦ 3. ТЦГ
15. Кількість т-РНК, що беруть участь у трансляції, дорівнює кількості:
1.Кодонів і-РНК, що шифрують амінокислоти 2. Молекул і-РНК
3 Генів, що входять до молекули ДНК 4. Білків, що синтезуються на рибосомах
16. Встановіть послідовність розташування нуклеотидів і-РНК при транскрипції з одним із ланцюгів ДНК: А-Г-Т-Ц-Г
1) У 2) Г 3) Ц 4) А 5) Ц
17. При реплікації молекули ДНК утворюється:
1) нитка, що розпалася на окремі фрагменти дочірніх молекул
2) молекула, що складається з двох нових ланцюгів ДНК
3) молекула, половина якої складається з нитки іРНК
4) дочірня молекула, що складається з одного старого та одного нового ланцюга ДНК
18. Матрицею для синтезу молекули іРНК при транскрипції служить:
1) вся молекула ДНК 2) повністю один із ланцюгів молекули ДНК
3) ділянка одного з ланцюгів ДНК
4) в одних випадках один із ланцюгів молекули ДНК, в інших – вся молекула ДНК.
19. Процес самоподвоєння молекули ДНК.
1. реплікація 2. репарація
3. реанкорнація
20. При біосинтезі білка в клітині енергія АТФ:
1) витрачається 2) запасається
3) не витрачається та не виділяється
21. У соматичних клітинах багатоклітинного організму:
1) різний набір генів та білків 2) однаковий набір генів та білків
3) однаковий набір генів, але різний набір білків
4) однаковий набір білків, але різний набір генів
22.. Один триплет ДНК несе інформацію про:
1) послідовності амінокислот у молекулі білка
2) ознакі організму 3) амінокислоті в молекулі синтезованого білка
4) склад молекули РНК
23. Який із процесів не відбувається в клітинах будь-якої будови та функції:
1) синтез білків 2) обмін речовин 3) мітоз 4) мейоз
24. Поняття «транскрипція» стосується процесу:
1) подвоєння ДНК 2) синтезу і-РНК на ДНК
3) переходу і-РНК на рибосоми 4) створення білкових молекул на полісомі
25. Ділянка молекули ДНК, що несе інформацію про одну молекулу білка - це:
1) ген 2) фенотип 3) геном 4) генотип
26. Транскрипція у еукаріотів відбувається в:
1) цитоплазмі 2) ендоплазматичній мембрані 3) лізосомах 4) ядрі
27.Синтез білка відбувається у:
1) гранулярному ендоплазматичному ретикулумі
2) гладкому ендоплазматичному ретикулумі 3) ядрі 4) лізосомах
28. Одна амінокислота кодується:
1) чотирма нуклеотидами 2) двома нуклеотидами
3) одним нуклеотидом 4) трьома нуклеотидами
29. Триплету нуклеотидів АТЦ у молекулі ДНК відповідатиме кодон молекули і-РНК:
1) ТАГ 2) УАГ 3) УТЦ 4) ЦАУ
30. Розділові знакигенетичного коду:
1.кодують певні білки 2. запускають синтез білка
3. припиняють синтез білка
31. Процес самоподвоєння молекули ДНК.
1.реплікація 2.репарація 3.реанкорнація
32. Функція і-РНК у процесі біосинтезу.
1.зберігання спадкової інформації 2.транспорт АК на рибосоми
3.подача інформації на рибосоми
33. Процес, коли т-РНК приносять амінокислоти рибосоми.
1.транскрипція 2.трансляція 3.трансформація
34. Рибосоми, що синтезують одну й ту саму білкову молекулу.
1.хромосома 2.полісома 3.мегахромосома
35. Процес, у якому амінокислоти утворюють білкову молекулу.
1.транскрипція 2.трансляція 3.трансформація
36. До реакцій матричного синтезу відносять...
1.реплікацію ДНК 2.транскрипцію, трансляцію 3.обидва відповіді правильні
37. Один триплет ДНК несе інформацію про:
1.Послідовності амінокислот у молекулі білка
2.Місце певної АК в білковому ланцюгу
3.Ознака конкретного організму
4.Амінокислоті, що включається в білковий ланцюг
38. У гені закодована інформація про:
1) будову білків, жирів та вуглеводів 2) первинній структурі білка
3) послідовності нуклеотидів у ДНК
4) послідовності амінокислот у 2-х і більше молекулах білків
39. Синтез іРНК починається з:
1) роз'єднання ДНК на дві нитки 2) взаємодії ферменту РНК - полімерази та гена
3) подвоєння гена 4) розпаду гена на нуклеотиди
40. Транскрипція відбувається:
1) у ядрі 2) на рибосомах 3) у цитоплазмі 4) на каналах гладкої ЕПС
41. Синтез білка не йде на рибосомах у:
1) збудника туберкульозу 2) бджоли 3) мухомору 4) бактеріофага
42. При трансляції матрицею для збирання поліпептидного ланцюга білка служать:
1) обидва ланцюжки ДНК 2) один з ланцюгів молекули ДНК
3) молекула іРНК 4) в одних випадках один з ланцюгів ДНК, в інших – молекула іРНК
Первинною структурою білків називається лінійний поліпептидний ланцюг з амінокислот, з'єднаних між собою пептидними зв'язками. Первинна структура – найпростіший рівень структурної організації білкової молекули. Високу стабільність їй надають ковалентні пептидні зв'язки між α-аміногрупою однієї амінокислоти та α-карбоксильною групою іншої амінокислоти.
Якщо в освіті пептидного зв'язку бере участь іміногрупа проліну або гідроксипроліну, то вона має інший вигляд

При утворенні пептидних зв'язків у клітинах спочатку активується карбоксильна група однієї амінокислоти, а потім вона з'єднується з аміногрупою іншою. Приблизно також проводять лабораторний синтез поліпептидів.
Пептидна зв'язок є фрагментом поліпептидного ланцюга, що повторюється. Вона має низку особливостей, які впливають не тільки на форму первинної структури, а й на найвищі рівні організації поліпептидного ланцюга:


· Копланарність - всі атоми, що входять до пептидної групи, знаходяться в одній площині;
· Здатність існувати у двох резонансних формах (кето-або енольной формі);
· транс-положення заступників стосовно С-N-зв'язку;
· Здатність до утворення водневих зв'язків, причому кожна з пептидних груп може утворювати два водневі зв'язки з іншими групами, у тому числі і пептидними.
Виняток становлять пептидні групи за участю аміногрупи проліну або гідроксипроліну. Вони здатні утворювати лише один водневий зв'язок (див. вище). Це позначається на формуванні вторинної структури білка. Поліпептидний ланцюг на ділянці, де знаходиться пролін або гідроксипролін, легко згинається, тому що не утримується, як завжди, другим водневим зв'язком.
схема утворення трипептиду:

Рівні просторової організації білків: вторинна структура білків: поняття про α-спіраль та β-складчастий шар. Третинна структура білків: поняття про нативний білок та денатурацію білка. Четвертична структура білків з прикладу будови гемоглобіну.
Вторинна структура білка.Під вторинною структурою білка розуміють спосіб укладання поліпептидного ланцюга у впорядковану структуру. По конфігурації виділяють такі елементи вторинної структури: α -спіраль та β -Складчастий шар.
Модель будови α-спіралі, що враховує всі властивості пептидного зв'язку, була розроблена Л. Полінгом та Р. Корі (1949 - 1951 рр.).
На малюнку 3, азображена схема α -спіралі, що дає уявлення про основні її параметри Поліпептидна ланцюг згортається в α -спіраль таким чином, що витки спіралі регулярні, тому спіральна конфігурація має гвинтову симетрію (рис. 3, б). на кожен виток α -спіралі припадає 3,6 амінокислотних залишків. Відстань між витками або крок спіралі становить 0,54 нм, кут підйому витка дорівнює 26 °. Формування та підтримка α -спіральної конфігурації відбувається за рахунок водневих зв'язків, що утворюються між пептидними групами кожного n-го та ( п+ 3)-го амінокислотних залишків. Хоча енергія водневих зв'язків мала, велика кількість їх призводить до значного енергетичного ефекту, внаслідок чого α -Спіральна конфігурація досить стійка. Бічні радикали амінокислотних залишків не беруть участь у підтримці α -спіральної конфігурації, тому всі амінокислотні залишки в α -Спіралі рівнозначні.
У природних білках існують лише правозакручені α -Спіралі.
β-Складчастий шар- Другий елемент вторинної структури. На відміну від α -спіралі β -Складчастий шар має лінійну, а не стрижневу форму (рис. 4). Лінійна структура утримується завдяки виникненню водневих зв'язків між пептидними угрупованнями, що стоять на різних ділянках поліпептидного ланцюга. Ці ділянки виявляються зближеними на відстань водневого зв'язку між - С = О та HN - групами (0,272 нм).

Мал. 4. Схематичне зображення β
-складчастого шару (стрілками вказано
 про напрямок поліпептидного ланцюга)
про напрямок поліпептидного ланцюга)
Мал. 3. Схема ( а) та модель ( б) α -спіралі
Вторинна структура білка визначається первинною. Амінокислотні залишки різною мірою здатні до утворення водневих зв'язків, це і впливає на утворення. α -спіралі або β -Шару. До спіралеутворюючих амінокислот відносяться аланін, глутамінова кислота, глутамін, лейцин, лізин, метіонін і гістидин. Якщо фрагмент білка складається головним чином із перерахованих вище амінокислотних залишків, то на даній ділянці сформується α -спіраль. Валін, ізолейцин, треонін, тирозин та фенілаланін сприяють утворенню β -Шарів поліпептидного ланцюга. Неупорядковані структури виникають на ділянках поліпептидного ланцюга, де сконцентровані такі амінокислотні залишки, як гліцин, серії, аспарагінова кислота, аспарагін, пролін.
У багатьох білках одночасно є і α -спіралі, та β -Шари. Частка спіральної конфігурації у різних білків різна. Так, м'язовий білок параміозин практично на 100% спіралізований; висока частка спіральної конфігурації у міоглобіну та гемоглобіну (75%). Навпаки, у трипсину та рибонуклеази значна частина поліпептидного ланцюга укладається в шаруваті β -структури. Білки опорних тканин - кератин (білок волосся), колаген (білок шкіри та сухожилля) - мають β -Конфігурацію поліпептидних ланцюгів.
Третинна структура білка.Третинна структура білка - це спосіб укладання поліпептидного ланцюга у просторі. Щоб білок придбав властиві йому функціональні властивості, поліпептидний ланцюг повинен певним чином згорнутися у просторі, сформувавши функціонально активну структуру. Така структура називається нативний. Незважаючи на величезну кількість теоретично можливих для окремого поліпептидного ланцюга просторових структур, згортання білка призводить до утворення єдиної нативної конфігурації.
Стабілізують третинну структуру білка взаємодії, що виникають між бічними радикалами амінокислотних залишків різних ділянок поліпептидного ланцюга. Ці взаємодії можна поділити на сильні та слабкі.
До сильних взаємодій відносяться ковалентні зв'язки між атомами сірки залишків цистеїну, що стоять у різних ділянках поліпептидного ланцюга. Інакше такі зв'язки називаються дисульфідними мостами; утворення дисульфідного мосту можна зобразити таким чином:
Крім ковалентних зв'язків третинна структура білкової молекули підтримується слабкими взаємодіями, які, своєю чергою, поділяються на полярні та неполярні.
До полярних взаємодій відносяться іонні та водневі зв'язки. Іонні взаємодії утворюються при контакті позитивно заряджених груп бічних радикалів лізину, аргініну, гістидину та негативно зарядженої СООН-групи аспарагінової та глутамінової кислот. Водневі зв'язки виникають між функціональними групами бічних радикалів амінокислотних залишків.
Неполярні або ван-дер-ваальсові взаємодії між вуглеводневими радикалами амінокислотних залишків сприяють формуванню гідрофобного ядра (Жирної краплі) всередині білкової глобули, т.к. Вуглеводневі радикали прагнуть уникнути зіткнення з водою. Чим більше у складі білка неполярних амінокислот, тим більшу роль у формуванні його третинної структури грають ван-дер-ваальсові зв'язки.
Численні зв'язки між бічними радикалами амінокислотних залишків визначають просторову конфігурацію білкової молекули (рис. 5).

Мал. 5. Типи зв'язків, що підтримують третинну структуру білка:
а- дисульфідний місток; б -іонний зв'язок; в, г -водневі зв'язки;
д -ван-дер-ваальсові зв'язки
Третинна структура окремо взятого білка унікальна, як унікальна та її первинна структура. Тільки правильне просторове укладання білка робить його активним. Різні порушення третинної структури призводять до зміни властивостей білка та втрати біологічної активності.
Четвертична структура білка.Білки з молекулярною масою понад 100 кДа 1 складаються, як правило, з кількох поліпептидних ланцюгів із порівняно невеликою молекулярною масою. Структура, що складається з певної кількості поліпептидних ланцюгів, що займають строго фіксоване положення відносно один одного, внаслідок чого білок має ту чи іншу активність, називається четвертинною структурою білка. Білок, що має четвертинну структуру, називається епімолекулоюабо мультимером , а складові його поліпептидні ланцюги - відповідно субодиницями або протомірами . Характерною властивістю білків з четвертинною структурою є те, що окрема субодиниця не має біологічної активності.
Стабілізація четвертинної структури білка відбувається за рахунок полярних взаємодій між бічними радикалами амінокислотних залишків, локалізованих на поверхні субодиниць. Такі взаємодії міцно утримують субодиниці як організованого комплексу. Ділянки субодиниць, у яких відбуваються взаємодії, називаються контактними майданчиками.
Класичним прикладом білка, що має четвертинну структуру, є гемоглобін. Молекула гемоглобіну з молекулярною масою 68 000 Так складається з чотирьох субодиниць двох різних типів. α і β / α -Субодиниця складається з 141 амінокислотного залишку, a β - із 146. Третинна структура α - І β -субодиниця подібна, як і їх молекулярна маса (17 000 Так). Кожна субодиниця містить простетичну групу - гем . Оскільки гем є і в інших білках (цитохроми, міоглобін), які вивчатимуться далі, хоча б коротко обговоримо структуру теми (рис. 6). Угруповання гема є складною копланарною циклічною системою, що складається з центрального атома, який утворює координаційні зв'язки з чотирма залишками піролу, з'єднаними метановими містками (= СН -). У гемоглобіні залізо зазвичай перебуває у стані окислення (2+).
Чотири субодиниці – дві α і дві β - з'єднуються в єдину структуру таким чином, що α -субодиниці контактують тільки з β -субодиницями і навпаки (рис. 7).

Мал. 6. Структура гему гемоглобіну

Мал. 7. Схематичне зображення четвертинної структури гемоглобіну:
Fe - гем гемоглобіну
Як видно з малюнка 7, одна молекула гемоглобіну здатна переносити 4 молекули кисню. І зв'язування, і звільнення кисню супроводжується конформаційними змінами структури α - І β -субодиниць гемоглобіну та їх взаємного розташування в епімолекулі Цей факт свідчить, що четвертинна структура білка перестав бути абсолютно жорсткої.
Подібна інформація.
Однією з особливостей білків є складна структурна організація. Всі білки мають первинну, вторинну і третинну структуру, а ті, які у своєму складі мають дві і більше ППЦ, мають і четвертинну структуру (НС).
Первинна структура білка (ПСБ) – це порядок чергування (послідовність) амінокислотних залишків у ППЦ.
Навіть однакові за своєю довжиною та амінокислотним складом білки можуть бути різними речовинами. Наприклад, з двох амінокислот можна скласти 2 різні дипептиди:
При числі амінокислот, що дорівнює 20, кількість можливих комбінацій дорівнює 210 18 . А якщо врахувати, що в ППЦ кожна амінокислота може зустрітися більше 1 разу, то кількість можливих варіантів важко підрахувати.
Визначення первинної структури білка (ПСБ).
ПСБ білків можна визначити за допомогою фенілтіогідантоїнового
методу
. Цей метод заснований на реакції взаємодії фенілізотіоціанату
(ФІТЦ) з α-АК. В результаті утворюється комплекс цих двох сполук ФІТЦ-АК
.
Наприклад, розглянемо пептид ![]() з метою визначення його ПСБ, тобто послідовності сполуки амінокислотних залишків.
з метою визначення його ПСБ, тобто послідовності сполуки амінокислотних залишків.
ФІТЦ взаємодіє з кінцевою амінокислотою (а). Утворюється комплекс ФТГ-а, його відокремлюють від суміші та визначають справжність амінокислоти. а. Наприклад, це – асн
![]() і т.д. Послідовно відокремлюють та ідентифікують всі інші амінокислоти. Це трудомісткий процес. Визначення ПСБ білка середнього розміру триває кілька місяців.
і т.д. Послідовно відокремлюють та ідентифікують всі інші амінокислоти. Це трудомісткий процес. Визначення ПСБ білка середнього розміру триває кілька місяців.
Пріоритет у розшифровці ПСБ належить Сенджеру(1953), який відкрив ПСБ інсуліну (Лауреат Нобелівської премії). Молекула інсуліну складається з 2х ППЦ – A та B.

А-ланцюг складається з 21 амінокислоти, ланцюг - з 30. Між собою ППЦ з'єднуються дисульфідними містками. Число білків, ПСБ яких визначено, на сьогодні досягає 1500. Навіть невеликі зміни первинної структури можуть суттєво змінити властивості білка. В еритроцитах здорових людей міститься HbA – при заміні в -ланцюзі HbA, у 6-му положенні глуна валвиникає тяжке захворювання серповидно-клітинна анемія, за якої діти, що народилися з цією аномалією, гинуть у ранньому віці. З іншого боку, можливі варіанти зміни ПСБ, які не позначаються на його фізико-хімічних та біологічних властивостях. Наприклад, HbC містить у 6-му положенні b-ланцюга замість глу – ліз, HbС майже не відрізняється за своїми властивостями від HbA, а люди, які мають в еритроцитах такий Hb, практично здорові.
Стабільність ПСБзабезпечується в основному міцними ковалентними пептидними зв'язками та, у другу чергу, дисульфідними зв'язками.
Вторинна структура білка (ВСБ).
ППЦ білків мають велику гнучкість і набувають певної просторової структури або конформацію. У білках розрізняють 2 рівні такої конформації – це ВСБ та третинна структура (ТББ).
ВСБ – це конфігурація ППЦ, тобто спосіб її укладання або скручування в якусь конформацію відповідно до програми, закладеної в ПСБ.
Відомі три основні типи ВСБ:
1) -спіраль;
2) b-структура(складчастий шар або складчастий листок);
3) безладний клубок.
-спіраль .
Її модель запропонована В. Полінг. Вона найімовірніша для глобулярних білків. Для будь-якої системи найбільш стійким є стан, що відповідає мінімуму вільної енергії. Для пептидів такий стан має місце, коли CO-і NH-групи з'єднуються між собою слабким водневим зв'язком. У a -спіралі NH-групи 1-го амінокислотного залишку взаємодіє з CO-групою 4-ї за рахунком амінокислотою. В результаті пептидний кістяк утворює спіраль, на кожен виток якої припадає 3,6 АК-залишку.
1 крок спіралі (1 виток) = 3,6 АК = 0,54 нм, кут підйому – 26°
Закручування ППЦ відбувається за годинниковою стрілкою, тобто у спіралі правий хід. Через кожні 5 витків (18 АК; 2,7 нм) конфігурація ППЦ повторюється.
Стабілізується ВСБнасамперед водневими зв'язками, і в другу – пептидними та дисульфідними. Водневі зв'язки в 10-100 разів слабші від звичайних хімічних зв'язків; однак за рахунок їх великої кількості вони забезпечують певну жорсткість та компактність ВСБ. Бічні R-ланцюги a-спіралі звернені до зовні та розташовані по різні боки від її осі.
b -структура .
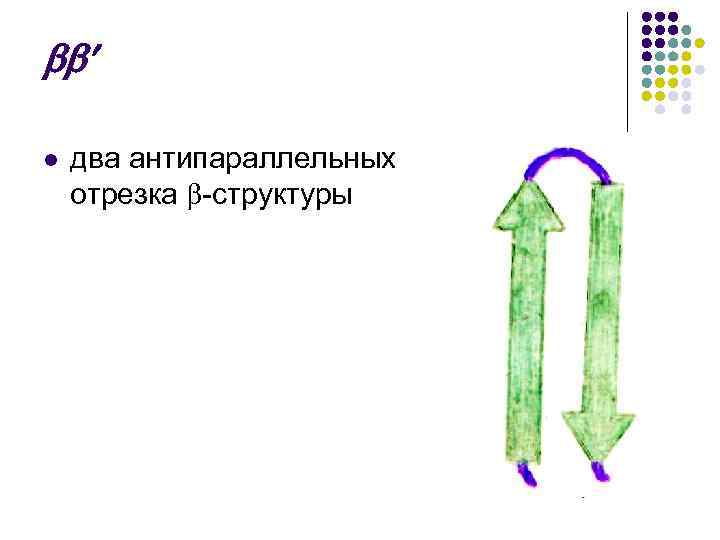
Це складчасті ділянки ППЦ, що формою нагадують листок, складений у гармошку. Шари ППЦ можуть бути паралельними, якщо обидва ланцюги починаються з N- або С-кінця.
Якщо суміжні ланцюги у шарі орієнтовані протилежними кінцями N–С та С–N, то вони називаються антипаралельними.

 паралельні
паралельні
![]() антипаралельні
антипаралельні
Утворення водневих зв'язків відбувається, як і в a-спіралі, між CO-і NH-групами.

 l l З'ясування структурної організації білків – одна з головних проблем сучасної біохімії Воно має важливе науково-практичне значення для розуміння величезного розмаїття функцій білків. з'єднуються між собою багато десятків і сотень амінокислот у білковій молекулі?
l l З'ясування структурної організації білків – одна з головних проблем сучасної біохімії Воно має важливе науково-практичне значення для розуміння величезного розмаїття функцій білків. з'єднуються між собою багато десятків і сотень амінокислот у білковій молекулі?
 Еміль Фішер Білки - складні поліпептиди, в яких окремі амінокислоти пов'язані один з одним пептидними (R-CO-NH-R) зв'язками, що виникають при взаємодії карбоксильних і аміногруп амінокислот
Еміль Фішер Білки - складні поліпептиди, в яких окремі амінокислоти пов'язані один з одним пептидними (R-CO-NH-R) зв'язками, що виникають при взаємодії карбоксильних і аміногруп амінокислот
 Експериментальні докази теорії l l l У природних білках порівняно мало вільних груп –СООН та –NH 2 У процесі гідролізу білка утворюється стехіометрична кількість титрованих груп – СООН та –NH 2 Під дією протеолітичних ферментів білки розщеплюються на строго визначені фрагменти (поліпептиди) Аналіз рентгенограм підтверджує поліпептидну структуру білка Істотне підтвердження поліпептидної теорії будови білка – можливість синтезу хімічними методами поліпептидів та білків з вже відомою будовою (інсуліну, лізоциму, рибонуклеа).
Експериментальні докази теорії l l l У природних білках порівняно мало вільних груп –СООН та –NH 2 У процесі гідролізу білка утворюється стехіометрична кількість титрованих груп – СООН та –NH 2 Під дією протеолітичних ферментів білки розщеплюються на строго визначені фрагменти (поліпептиди) Аналіз рентгенограм підтверджує поліпептидну структуру білка Істотне підтвердження поліпептидної теорії будови білка – можливість синтезу хімічними методами поліпептидів та білків з вже відомою будовою (інсуліну, лізоциму, рибонуклеа).

 Структурні особливості пептидного ланцюга l l Пептидний зв'язок приблизно на 10% коротший від зв'язку –С–N– і має характер «частково подвійного» зв'язку –С=N– Л. Полінг та Р. Корі запропонували у 1948–1955 рр. пояснювати особливу природу зв'язку С–N «резонансом» між двома формами
Структурні особливості пептидного ланцюга l l Пептидний зв'язок приблизно на 10% коротший від зв'язку –С–N– і має характер «частково подвійного» зв'язку –С=N– Л. Полінг та Р. Корі запропонували у 1948–1955 рр. пояснювати особливу природу зв'язку С–N «резонансом» між двома формами
 Структурні особливості пептидного ланцюга l l l Іншими словами, у білках і пептидах зв'язок С–N є частково кратним через взаємодію неподіленої пари електронів атома азоту з електронною системою карбонільної групи, що призводить до утрудненого обертання навколо зв'язку С–N Зазвичай пептидний зв'язок має транс- конфігурацію, що вигідніше цис-конфігурації на 2, 6 ккал/моль (10, 878 к. Дж/моль), т. к. зближення -вуглецевих атомів у цисконфігурації викликає просторові труднощі про
Структурні особливості пептидного ланцюга l l l Іншими словами, у білках і пептидах зв'язок С–N є частково кратним через взаємодію неподіленої пари електронів атома азоту з електронною системою карбонільної групи, що призводить до утрудненого обертання навколо зв'язку С–N Зазвичай пептидний зв'язок має транс- конфігурацію, що вигідніше цис-конфігурації на 2, 6 ккал/моль (10, 878 к. Дж/моль), т. к. зближення -вуглецевих атомів у цисконфігурації викликає просторові труднощі про
 Структурні особливості пептидного ланцюга l l l Повороти можливі навколо двох простих зв'язків (N–С і С–C 1), що примикають до асиметричного атома зазвичай = 180 - транс-форма)
Структурні особливості пептидного ланцюга l l l Повороти можливі навколо двох простих зв'язків (N–С і С–C 1), що примикають до асиметричного атома зазвичай = 180 - транс-форма)
 Структурні особливості пептидного ланцюга l l В силу взаємодії між заступниками в пептидному ланцюгу кути і не можуть приймати будь-які значення – для них дозволеними виявляються лише деякі дискретні області, що відповідають енергетично вигідним конформаціям пептидного ланцюга Кути і взаємопов'язані, зміна одного з них тягне зміну другого
Структурні особливості пептидного ланцюга l l В силу взаємодії між заступниками в пептидному ланцюгу кути і не можуть приймати будь-які значення – для них дозволеними виявляються лише деякі дискретні області, що відповідають енергетично вигідним конформаціям пептидного ланцюга Кути і взаємопов'язані, зміна одного з них тягне зміну другого

 l К. У. Ліндерстрем-Ланг запропонував розрізняти 4 рівня організації білкових молекул – первинну, вторинну, третинну та четвертинну структури. Хоча ці категорії певною мірою застаріли, ними поки що продовжують користуватися.
l К. У. Ліндерстрем-Ланг запропонував розрізняти 4 рівня організації білкових молекул – первинну, вторинну, третинну та четвертинну структури. Хоча ці категорії певною мірою застаріли, ними поки що продовжують користуватися.

 Первинна структура білка l l l послідовність амінокислотних залишків у поліпептидному ланцюгу кодується структурним геном даного білка і містить у собі все необхідне для самоорганізації його просторової структури Всі білки різняться за своєю первинною структурою Потенційно можлива кількість таких структур необмежена кількість різних типів білків у всіх видів живих організмів становить величину порядку 1010-1012
Первинна структура білка l l l послідовність амінокислотних залишків у поліпептидному ланцюгу кодується структурним геном даного білка і містить у собі все необхідне для самоорганізації його просторової структури Всі білки різняться за своєю первинною структурою Потенційно можлива кількість таких структур необмежена кількість різних типів білків у всіх видів живих організмів становить величину порядку 1010-1012
 l l l Пізнання біологічної функції і, зокрема, молекулярного механізму фізіологічної дії білка неможливе без детального знання його будови. Дослідження первинної структури «мутагенних» білків дозволяє на молекулярному рівні з'ясувати характер спадкових хвороб.
l l l Пізнання біологічної функції і, зокрема, молекулярного механізму фізіологічної дії білка неможливе без детального знання його будови. Дослідження первинної структури «мутагенних» білків дозволяє на молекулярному рівні з'ясувати характер спадкових хвороб.
 Методи визначення первинної структури l l Підготовка білка до аналізу первинної структури покликана звести до мінімуму вплив інших, більш високих рівнів його організації Іншими словами, об'єктом аналізу має бути невпорядкований білковий ланцюг без будь-яких поперечних ковалентних зв'язків (наприклад, дисульфідних) так, щоб усі його ланки, всі пептидні зв'язки були однаково доступні дії як хімічних реагентів, так і ферментів
Методи визначення первинної структури l l Підготовка білка до аналізу первинної структури покликана звести до мінімуму вплив інших, більш високих рівнів його організації Іншими словами, об'єктом аналізу має бути невпорядкований білковий ланцюг без будь-яких поперечних ковалентних зв'язків (наприклад, дисульфідних) так, щоб усі його ланки, всі пептидні зв'язки були однаково доступні дії як хімічних реагентів, так і ферментів
 l l Білок повинен бути перш за все підданий глибокій денатурації і втратити четвертинну, третинну і по можливості вторинну структуру.
l l Білок повинен бути перш за все підданий глибокій денатурації і втратити четвертинну, третинну і по можливості вторинну структуру.
 l Щоб уникнути замикання дисульфідних зв'язків, знову сульфгідрильні групи блокують надлишком йодоцтової кислоти: R–SН + I–СН 2–СООН R–S–СН 2–СООН + НI
l Щоб уникнути замикання дисульфідних зв'язків, знову сульфгідрильні групи блокують надлишком йодоцтової кислоти: R–SН + I–СН 2–СООН R–S–СН 2–СООН + НI
 Потім проводять селективний гідроліз денатурованого білка для отримання системи пептидів, що перекриваються. Гідроліз проводять ферментами: l l l трипсином (зв'язки, утворені карбоксильними групами арг або ліз), хімотрипсином (зв'язки, утворені групами -СООН ароматичних амінокислот) та ін; хімічними реагентами: бромціаном Br - С N (зв'язки, утворені групами - СООН мет) та ін.
Потім проводять селективний гідроліз денатурованого білка для отримання системи пептидів, що перекриваються. Гідроліз проводять ферментами: l l l трипсином (зв'язки, утворені карбоксильними групами арг або ліз), хімотрипсином (зв'язки, утворені групами -СООН ароматичних амінокислот) та ін; хімічними реагентами: бромціаном Br - С N (зв'язки, утворені групами - СООН мет) та ін.
 l l l Наступний етап – фракціонування отриманих пептидів – здійснюється методами хроматографії Нарешті, останній етап – визначення первинної структури l починається з визначення його молекулярної маси, амінокислотного складу, N- і Кінцевих амінокислотних залишків. іншого агента та отримують другий, відмінний від першого набір пептидних фрагментів, які поділяють та аналізують для з'ясування порядку розташування амінокислот у білку
l l l Наступний етап – фракціонування отриманих пептидів – здійснюється методами хроматографії Нарешті, останній етап – визначення первинної структури l починається з визначення його молекулярної маси, амінокислотного складу, N- і Кінцевих амінокислотних залишків. іншого агента та отримують другий, відмінний від першого набір пептидних фрагментів, які поділяють та аналізують для з'ясування порядку розташування амінокислот у білку

 Метод Ф. Сенджера l У 1945 р. англійський біохімік Ф. Сенджер запропонував один із перших методів визначення N-кінцевих амінокислот
Метод Ф. Сенджера l У 1945 р. англійський біохімік Ф. Сенджер запропонував один із перших методів визначення N-кінцевих амінокислот
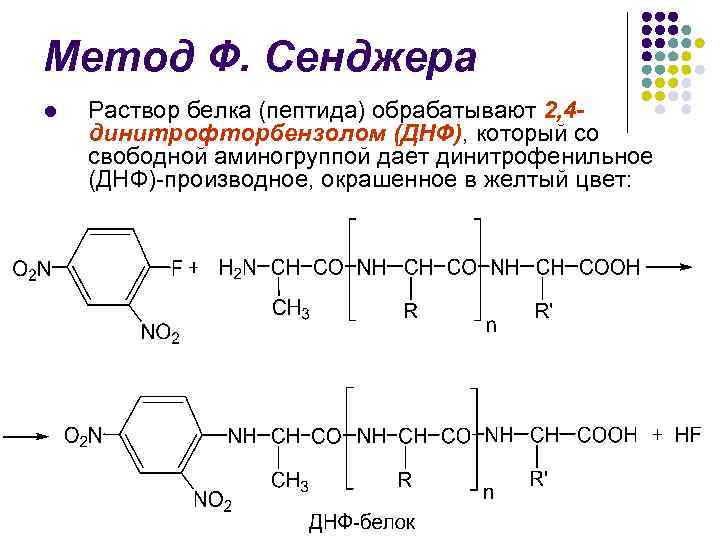 Метод Ф. Сенджера l Розчин білка (пептиду) обробляють 2, 4 динітрофторбензолом (ДНФ), який із вільною аміногрупою дає динітрофенільне (ДНФ)-похідне, забарвлене у жовтий колір:
Метод Ф. Сенджера l Розчин білка (пептиду) обробляють 2, 4 динітрофторбензолом (ДНФ), який із вільною аміногрупою дає динітрофенільне (ДНФ)-похідне, забарвлене у жовтий колір:
 Метод Ф. Сенджера l l Наступний кислотний гідроліз (5, 7 н. НСl) призводить до розриву пептидних зв'язків та утворення ДНФ-похідного Nкінцевої амінокислоти.
Метод Ф. Сенджера l l Наступний кислотний гідроліз (5, 7 н. НСl) призводить до розриву пептидних зв'язків та утворення ДНФ-похідного Nкінцевої амінокислоти.
 Дансильний метод l Розроблений в 1963 р. англійськими біохіміками В. Греєм і Б. Хартлі
Дансильний метод l Розроблений в 1963 р. англійськими біохіміками В. Греєм і Б. Хартлі
 Дансильний метод l Перша стадія – реакція дансилхлориду (1-диметиламінонафталін-5-сульфохлориду) з непротонованою -аміногрупою пептиду (або білка) з утворенням дансилпептиду (ДНС-пептиду)
Дансильний метод l Перша стадія – реакція дансилхлориду (1-диметиламінонафталін-5-сульфохлориду) з непротонованою -аміногрупою пептиду (або білка) з утворенням дансилпептиду (ДНС-пептиду)

 Дансильний метод Отриманий ДНС-білок піддається гідролізу в присутності 5, 7 н. НСl, при 105 С протягом 12-16 год, що відщепилася ДНС-амінокислоту ідентифікують за флуоресценцією в УФ-променях
Дансильний метод Отриманий ДНС-білок піддається гідролізу в присутності 5, 7 н. НСl, при 105 С протягом 12-16 год, що відщепилася ДНС-амінокислоту ідентифікують за флуоресценцією в УФ-променях


 Метод С. Акабори l l При нагріванні пептиду (або білка) з безводним гідразином при 100-120 С пептидні зв'язки гідролізуються з утворенням амінокислот гідразидів С-кінцева амінокислота залишається у вигляді вільної амінокислоти і може бути виділена з суміші та ідентифікована:
Метод С. Акабори l l При нагріванні пептиду (або білка) з безводним гідразином при 100-120 С пептидні зв'язки гідролізуються з утворенням амінокислот гідразидів С-кінцева амінокислота залишається у вигляді вільної амінокислоти і може бути виділена з суміші та ідентифікована:

 Метод П. Едмана l Метод деградації поліпептидного ланцюга за допомогою фенілізотиоціонату (ФІТЦ) розроблений шведським хіміком П. В. Едманом у 1950-56 рр. l Метод Едмана дозволяє послідовно відщеплювати N-кінцеві амінокислотні залишки у вигляді фенілтіогідантоїнів (ФТГ) l Кожен цикл деградації включає 3 стадії
Метод П. Едмана l Метод деградації поліпептидного ланцюга за допомогою фенілізотиоціонату (ФІТЦ) розроблений шведським хіміком П. В. Едманом у 1950-56 рр. l Метод Едмана дозволяє послідовно відщеплювати N-кінцеві амінокислотні залишки у вигляді фенілтіогідантоїнів (ФТГ) l Кожен цикл деградації включає 3 стадії
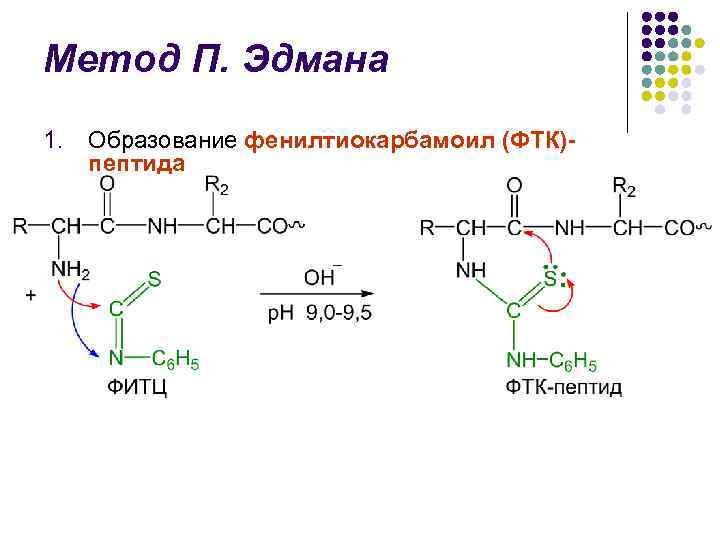



 Метод П. Едмана l l l Ідентифікація відщеплених ФТГ є визначальною в методі Едмана Протягом тривалого часу використовували хроматографію на папері, потім мікротонкошарова хроматографію на силікагелі і поліаміді, рідинну і газо-рідинну хроматографії Великим досягненням в області структурних досліджень. Едманом та Дж. Беггом секвенатора (від англ. sequence – послідовність) – приладу, який з високою ефективністю здійснює послідовне автоматичне відщеплення N-кінцевих амінокислотних залишків за методом Едмана
Метод П. Едмана l l l Ідентифікація відщеплених ФТГ є визначальною в методі Едмана Протягом тривалого часу використовували хроматографію на папері, потім мікротонкошарова хроматографію на силікагелі і поліаміді, рідинну і газо-рідинну хроматографії Великим досягненням в області структурних досліджень. Едманом та Дж. Беггом секвенатора (від англ. sequence – послідовність) – приладу, який з високою ефективністю здійснює послідовне автоматичне відщеплення N-кінцевих амінокислотних залишків за методом Едмана
 l Для визначення структури пептидів та білків можна застосовувати: l l ферменти, що каталізують відщеплення N- та С-кінцевих амінокислотних залишків – аміної карбоксипептидази фізико-хімічні методи, зокрема, мас-спектрометрію
l Для визначення структури пептидів та білків можна застосовувати: l l ферменти, що каталізують відщеплення N- та С-кінцевих амінокислотних залишків – аміної карбоксипептидази фізико-хімічні методи, зокрема, мас-спектрометрію
 Аналіз даних по первинній структурі дозволяє зробити наступні загальні висновки Стабільність первинної структури забезпечується в основному пептидними зв'язками; можлива участь і невеликої кількості дисульфідних зв'язків У поліпептидному ланцюгу можуть бути виявлені різноманітні комбінації амінокислот У поліпептидах були відкриті всі можливі дипептиди
Аналіз даних по первинній структурі дозволяє зробити наступні загальні висновки Стабільність первинної структури забезпечується в основному пептидними зв'язками; можлива участь і невеликої кількості дисульфідних зв'язків У поліпептидному ланцюгу можуть бути виявлені різноманітні комбінації амінокислот У поліпептидах були відкриті всі можливі дипептиди
 Аналіз даних по первинній структурі дозволяє зробити наступні загальні висновки: l l Кожен індивідуальний гомогенний білок характеризується унікальною первинною структурою; Часто заміни амінокислот призводять не тільки до структурних перебудов, а й до змін фізикохімічних властивостей і біологічних функцій. Цей принцип структурної подоби найбільш типовий для низки протеолітичних ферментів – трипсину, хімотрипсину та ін.
Аналіз даних по первинній структурі дозволяє зробити наступні загальні висновки: l l Кожен індивідуальний гомогенний білок характеризується унікальною первинною структурою; Часто заміни амінокислот призводять не тільки до структурних перебудов, а й до змін фізикохімічних властивостей і біологічних функцій. Цей принцип структурної подоби найбільш типовий для низки протеолітичних ферментів – трипсину, хімотрипсину та ін.

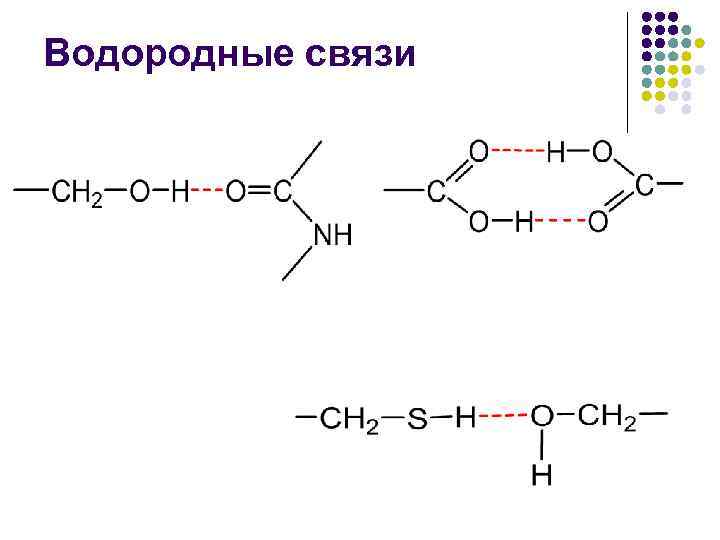
 Водневі зв'язки У неполярному оточенні енергія водневого зв'язку –СО НN– становить близько 16, 7 к. Дж/моль, а підвищення полярності середовища знижує цю енергію
Водневі зв'язки У неполярному оточенні енергія водневого зв'язку –СО НN– становить близько 16, 7 к. Дж/моль, а підвищення полярності середовища знижує цю енергію
 Гідрофобні взаємодії мають ентропійну природу Пов'язані з тим, що неполярні заступники виштовхуються з води і прагнуть обмежити свій контакт з нею Навпаки, вода прагне відновити свій структурований стан і як би примусово групує заступники в кластери, що володіють мінімумом енергії. залишків
Гідрофобні взаємодії мають ентропійну природу Пов'язані з тим, що неполярні заступники виштовхуються з води і прагнуть обмежити свій контакт з нею Навпаки, вода прагне відновити свій структурований стан і як би примусово групує заступники в кластери, що володіють мінімумом енергії. залишків

 Ван-дер-ваальсові взаємодії l l Складаються з дисперсійних сил тяжіння атомів і сил взаємного відштовхування їх електронних оболонок Енергетичний внесок кожного контакту невеликий (
Ван-дер-ваальсові взаємодії l l Складаються з дисперсійних сил тяжіння атомів і сил взаємного відштовхування їх електронних оболонок Енергетичний внесок кожного контакту невеликий (
 Іонні, або електростатичні, взаємодії l Взаємодії заряджених груп l l Взаємодії іоногенних груп, що утворюють сольові зв'язки Енергія сольових зв'язків у гідрофобному оточенні може досягати 41, 9 к. Дж/моль, але їх кількість порівняно невелика. аналогічні іон-дипольні та диполь-дипольні взаємодії
Іонні, або електростатичні, взаємодії l Взаємодії заряджених груп l l Взаємодії іоногенних груп, що утворюють сольові зв'язки Енергія сольових зв'язків у гідрофобному оточенні може досягати 41, 9 к. Дж/моль, але їх кількість порівняно невелика. аналогічні іон-дипольні та диполь-дипольні взаємодії
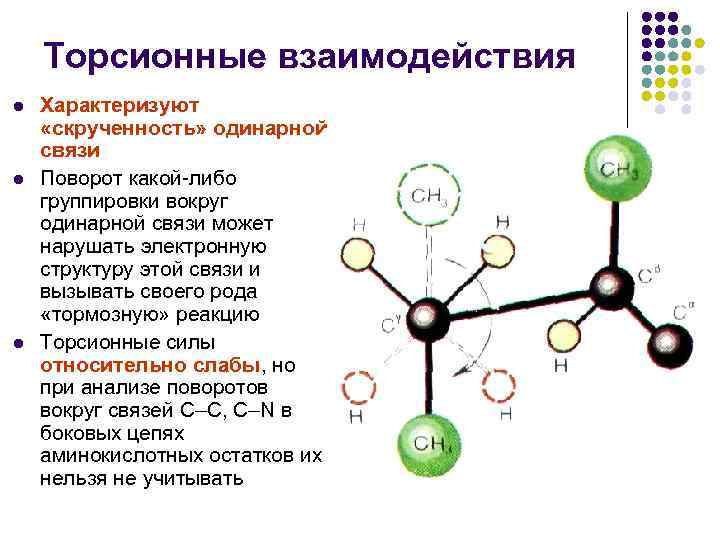 Торсійні взаємодії l l l Характеризують «скрученість» одинарного зв'язку Поворот будь-якого угруповання навколо одинарного зв'язку може порушувати електронну структуру цього зв'язку і викликати свого роду «гальмівну» реакцію Торсійні сили відносно слабкі, але при аналізі поворотів навколо зв'язків С–С, С–N бічних ланцюгах амінокислотних залишків їх не можна не враховувати
Торсійні взаємодії l l l Характеризують «скрученість» одинарного зв'язку Поворот будь-якого угруповання навколо одинарного зв'язку може порушувати електронну структуру цього зв'язку і викликати свого роду «гальмівну» реакцію Торсійні сили відносно слабкі, але при аналізі поворотів навколо зв'язків С–С, С–N бічних ланцюгах амінокислотних залишків їх не можна не враховувати

 Вторинна структура білка… l l просторове розташування окремих ділянок поліпептидного ланцюга без урахування типу та конформації бічних радикалів амінокислот Утворюється за рахунок водневих зв'язків між пептидними групами як одного ланцюга, так і різних ланцюгів
Вторинна структура білка… l l просторове розташування окремих ділянок поліпептидного ланцюга без урахування типу та конформації бічних радикалів амінокислот Утворюється за рахунок водневих зв'язків між пептидними групами як одного ланцюга, так і різних ланцюгів
 Вторинна структура білка l l l Будь-яка ділянка молекули білка має вторинну структуру Іноді розглядають як вторинну структуру лише періодичні її елементи: -спіраль і -структуру Однак у білках зустрічаються ділянки, які укладені цілком певним способом, хоча їхня просторова структура не містить будь-якого періодично повторюваного, регулярного мотиву. Проте до них цілком придатне поняття вторинної структури Розрізняють 2 види вторинних структур: регулярні та нерегулярні Поняття вторинної структури відноситься не до всієї білкової молекули в цілому, а до окремих більш-менш протяжних ділянок її поліпептидного ланцюга
Вторинна структура білка l l l Будь-яка ділянка молекули білка має вторинну структуру Іноді розглядають як вторинну структуру лише періодичні її елементи: -спіраль і -структуру Однак у білках зустрічаються ділянки, які укладені цілком певним способом, хоча їхня просторова структура не містить будь-якого періодично повторюваного, регулярного мотиву. Проте до них цілком придатне поняття вторинної структури Розрізняють 2 види вторинних структур: регулярні та нерегулярні Поняття вторинної структури відноситься не до всієї білкової молекули в цілому, а до окремих більш-менш протяжних ділянок її поліпептидного ланцюга
 Вторинна структура білка l Взаємодії, яким належить найважливіша роль у формуванні вторинної структури, визначаються: l l l особливостями пептидного зв'язку стеричними обмеженнями (- і -кути) Основну роль відіграють водневі зв'язки між пептидними угрупованнями, що періодично повторюються в ланцюзі
Вторинна структура білка l Взаємодії, яким належить найважливіша роль у формуванні вторинної структури, визначаються: l l l особливостями пептидного зв'язку стеричними обмеженнями (- і -кути) Основну роль відіграють водневі зв'язки між пептидними угрупованнями, що періодично повторюються в ланцюзі
 Стабілізація вторинної структури за рахунок водневого зв'язку l l Вплив навколишнього білок води Вода здатна ефективно конкурувати за утворення водневих зв'язків: Вплив води знижується при формуванні компактної просторової структури білка, зростанні вмісту пептидних зв'язків, підвищенні ймовірності їх взаємодії Таким чином, стабільність вторинної структури залежить від її у компактну третинну структуру
Стабілізація вторинної структури за рахунок водневого зв'язку l l Вплив навколишнього білок води Вода здатна ефективно конкурувати за утворення водневих зв'язків: Вплив води знижується при формуванні компактної просторової структури білка, зростанні вмісту пептидних зв'язків, підвищенні ймовірності їх взаємодії Таким чином, стабільність вторинної структури залежить від її у компактну третинну структуру

 -Спіраль l У 50-х роках ХХ ст. Л. Полінг і Р. Корі, ґрунтуючись на даних про структуру кристалів амінокислот і простих пептидів, розглянули можливі періодичні конформації поліпептидного ланцюга і дійшли висновку, що найбільш ймовірна структура, названа ними -спіраллю
-Спіраль l У 50-х роках ХХ ст. Л. Полінг і Р. Корі, ґрунтуючись на даних про структуру кристалів амінокислот і простих пептидів, розглянули можливі періодичні конформації поліпептидного ланцюга і дійшли висновку, що найбільш ймовірна структура, названа ними -спіраллю
 -Спіраль В основу її вибору були покладені наступні критерії: 1. Утворення щільноупакованої компактної структури без порожнеч і перекриття атомів 2. Максимальна насиченість структури водневими зв'язками з тією умовою, щоб їх геометрія була близька до лінійної 3. Дотримання міжатомних відстаней і кутів і простим пептидам l
-Спіраль В основу її вибору були покладені наступні критерії: 1. Утворення щільноупакованої компактної структури без порожнеч і перекриття атомів 2. Максимальна насиченість структури водневими зв'язками з тією умовою, щоб їх геометрія була близька до лінійної 3. Дотримання міжатомних відстаней і кутів і простим пептидам l
 -Спіраль l З дотриманням цих умов можна побудувати як праву, так і ліву спіралі, проте права -спіраль виявляється енергетично дещо вигідніше лівої, якщо пептидна ланцюг утворена L-амінокислотами
-Спіраль l З дотриманням цих умов можна побудувати як праву, так і ліву спіралі, проте права -спіраль виявляється енергетично дещо вигідніше лівої, якщо пептидна ланцюг утворена L-амінокислотами
 -Спіраль l l Спіральна структура виникає, коли у всіх ланках поліпептидного ланцюга кути повороту навколо простих зв'язків (і) мають однакову величину і знак (близькі до 60), що і призводить до поступового закручування ланцюга При цьому радикали залишків амінокислот виявляються на периферії утвореного спіраллю циліндра і можуть, залежно від їх характеру, забезпечувати гідрофобну або гідрофільну природу циліндричної поверхні
-Спіраль l l Спіральна структура виникає, коли у всіх ланках поліпептидного ланцюга кути повороту навколо простих зв'язків (і) мають однакову величину і знак (близькі до 60), що і призводить до поступового закручування ланцюга При цьому радикали залишків амінокислот виявляються на периферії утвореного спіраллю циліндра і можуть, залежно від їх характеру, забезпечувати гідрофобну або гідрофільну природу циліндричної поверхні
 -Геометричні параметри спіралі: l l l l радіус r = 2,3 Å (0,23 нм) висота спіралі (зміщення) на 1 залишок d = 1,5 Å крок спіралі (період ідентичності) P = 5,4 Å 1 виток спіралі, форма 3, 6 амінокислотних залишків, усі – Зв’язки С=О спрямовані вперед, до С-кінця, а групи –N–H – назад.У спіралі кожна група –NH з’єднана водневим зв’язком з групою –СО четвертого амінокислотного залишку. з нього (5 1 зв'язок)
-Геометричні параметри спіралі: l l l l радіус r = 2,3 Å (0,23 нм) висота спіралі (зміщення) на 1 залишок d = 1,5 Å крок спіралі (період ідентичності) P = 5,4 Å 1 виток спіралі, форма 3, 6 амінокислотних залишків, усі – Зв’язки С=О спрямовані вперед, до С-кінця, а групи –N–H – назад.У спіралі кожна група –NH з’єднана водневим зв’язком з групою –СО четвертого амінокислотного залишку. з нього (5 1 зв'язок)
 -Спіраль l l Довжина -спіральних ділянок в глобулярних білках відносно невелика (5 - 15 амінокислотних залишків, 3 - 4 витка спіралі), в фібрилярних білках - набагато протяжніше Іноді спостерігаються злами -спіралі, зазвичай в місцях включення залишків про, що переривають системи. При цьому вісь спіралі відхиляється на 20-30
-Спіраль l l Довжина -спіральних ділянок в глобулярних білках відносно невелика (5 - 15 амінокислотних залишків, 3 - 4 витка спіралі), в фібрилярних білках - набагато протяжніше Іноді спостерігаються злами -спіралі, зазвичай в місцях включення залишків про, що переривають системи. При цьому вісь спіралі відхиляється на 20-30

 -Структура l l Запропонована У. Т. Астбері в 1941 р. на підставі рентгеноструктурних досліджень -кератину Через 10 років Л. Полінг та Р. Корі встановили, що структура, або «складчастий лист», – це стабілізований міжланцюжковими водневими зв'язками асоціат витягнутих, зигзаг пептидних ланцюгів
-Структура l l Запропонована У. Т. Астбері в 1941 р. на підставі рентгеноструктурних досліджень -кератину Через 10 років Л. Полінг та Р. Корі встановили, що структура, або «складчастий лист», – це стабілізований міжланцюжковими водневими зв'язками асоціат витягнутих, зигзаг пептидних ланцюгів




 -Структура l l Число амінокислотних залишків у відрізку пептидного ланцюга, що утворює -структуру, зазвичай від 3 до 8 Протяжна -структура, так званий -шар, або -складчастий лист найчастіше складається з 2-6 ланцюгів, іноді до 10
-Структура l l Число амінокислотних залишків у відрізку пептидного ланцюга, що утворює -структуру, зазвичай від 3 до 8 Протяжна -структура, так званий -шар, або -складчастий лист найчастіше складається з 2-6 ланцюгів, іноді до 10
 -Структура l l l Бічні групи амінокислотних залишків при утворенні структури виявляються по різні боки її поверхні Сама ж поверхня має складчасту форму, причому складки задані -вуглецевими атомами Бічні групи, що відходять від них, утворюють гребені Це дозволяє формувати досить протяжні поверхні, насичені однотипними (наприклад, гідрофобними) бічними радикалами Гідрофобні поверхні -складчастого шару, взаємодіючи між собою або з гідрофобними гребенями -спіралей, беруть участь у побудові внутрішньомолекулярних гідрофобних ядер, що стабілізують просторову структуру білка
-Структура l l l Бічні групи амінокислотних залишків при утворенні структури виявляються по різні боки її поверхні Сама ж поверхня має складчасту форму, причому складки задані -вуглецевими атомами Бічні групи, що відходять від них, утворюють гребені Це дозволяє формувати досить протяжні поверхні, насичені однотипними (наприклад, гідрофобними) бічними радикалами Гідрофобні поверхні -складчастого шару, взаємодіючи між собою або з гідрофобними гребенями -спіралей, беруть участь у побудові внутрішньомолекулярних гідрофобних ядер, що стабілізують просторову структуру білка

 -Вигин l l Як -спіраль, так і -структура зазвичай представлені в глобулярних білках порівняно короткими відрізками, тому значна частина вторинної структури білка припадає на різного роду петлі, що дозволяють змінити напрям пептидного ланцюга Найбільш економний структурний елемент, що дозволяє повернути поліпептид на 180 . 3 пептидні угруповання, - -вигин Стабілізується одним водневим зв'язком Практично завжди виявляється на поверхні білкової глобули, тому нерідко відіграє істотну роль у її взаємодії з іншими молекулами
-Вигин l l Як -спіраль, так і -структура зазвичай представлені в глобулярних білках порівняно короткими відрізками, тому значна частина вторинної структури білка припадає на різного роду петлі, що дозволяють змінити напрям пептидного ланцюга Найбільш економний структурний елемент, що дозволяє повернути поліпептид на 180 . 3 пептидні угруповання, - -вигин Стабілізується одним водневим зв'язком Практично завжди виявляється на поверхні білкової глобули, тому нерідко відіграє істотну роль у її взаємодії з іншими молекулами


 l За результатами рентгеноструктурного аналізу виявлено, що спосіб укладання молекули білка визначається його амінокислотною послідовністю l l l освіті -спіралі сприяють ала, лей, глу освіті -структури - мет, вал, або вигину ланцюга - глі, про, асн Якщо з 6 згрупованих залишків амінокислот 4 сприяють утворенню спіралі, то дана група – центр спіралізації Якщо 3 залишки з 5 згрупованих сприяють утворенню -структури, то дане угруповання - затравка для
l За результатами рентгеноструктурного аналізу виявлено, що спосіб укладання молекули білка визначається його амінокислотною послідовністю l l l освіті -спіралі сприяють ала, лей, глу освіті -структури - мет, вал, або вигину ланцюга - глі, про, асн Якщо з 6 згрупованих залишків амінокислот 4 сприяють утворенню спіралі, то дана група – центр спіралізації Якщо 3 залишки з 5 згрупованих сприяють утворенню -структури, то дане угруповання - затравка для

 Надвторинна (надвторинна) структура білків l l Цей рівень організації білкової молекули пов'язаний з наявністю ансамблів взаємодіючих між собою вторинних структур.
Надвторинна (надвторинна) структура білків l l Цей рівень організації білкової молекули пов'язаний з наявністю ансамблів взаємодіючих між собою вторинних структур.

 Суперспіралізована -спіраль l l Утворюється при скручуванні декількох -спіралей у протофібрили (-кератин), які об'єднуються в мікрофібрили -Спіралі утримуються силами Ван-дер-Ваальса
Суперспіралізована -спіраль l l Утворюється при скручуванні декількох -спіралей у протофібрили (-кератин), які об'єднуються в мікрофібрили -Спіралі утримуються силами Ван-дер-Ваальса

 х l ланка, що складається з двох паралельних шарів з зчленуванням між ними у вигляді: l невпорядкованого клубка - с
х l ланка, що складається з двох паралельних шарів з зчленуванням між ними у вигляді: l невпорядкованого клубка - с
 х l ланка, що складається з двох паралельних -шарів з зчленуванням між ними у вигляді: l l -спіралі - укладання ланцюга по Россману Два послідовно з'єднаних ділянки утворюють структуру «грецький орнамент» -
х l ланка, що складається з двох паралельних -шарів з зчленуванням між ними у вигляді: l l -спіралі - укладання ланцюга по Россману Два послідовно з'єднаних ділянки утворюють структуру «грецький орнамент» -
 х l ланка, що складається з двох паралельних -шарів з зчленуванням між ними у вигляді: l -структури - - -зигзаг, «грецький ключ»
х l ланка, що складається з двох паралельних -шарів з зчленуванням між ними у вигляді: l -структури - - -зигзаг, «грецький ключ»

 Домени l l Багато білків містять відносно слабко взаємодіючі між собою ділянки – домени Домени (від фр. domaine – володіння) – структурно та функціонально відокремлені області (субобласті) молекули, з'єднані один з одним короткими ділянками поліпептидного ланцюга, які називаються шарнірними ділянками Функціональні домени можуть складатися з одного або декількох структурних доменів У ряду ферментів у поглибленнях між доменами розташовується активний центр
Домени l l Багато білків містять відносно слабко взаємодіючі між собою ділянки – домени Домени (від фр. domaine – володіння) – структурно та функціонально відокремлені області (субобласті) молекули, з'єднані один з одним короткими ділянками поліпептидного ланцюга, які називаються шарнірними ділянками Функціональні домени можуть складатися з одного або декількох структурних доменів У ряду ферментів у поглибленнях між доменами розташовується активний центр



 3. + -Білки l є ділянки, повністю побудовані з-спіралей, і ділянки, повністю побудовані з антипаралельних-шарів Інсулін
3. + -Білки l є ділянки, повністю побудовані з-спіралей, і ділянки, повністю побудовані з антипаралельних-шарів Інсулін

 4. / -Білки l l -Спіралі та -структури чергуються по ходу ланцюга Більшість -структур (паралельні) виявляються локалізованими в центральній частині молекули, де ці структури згинаються у вигляді пропелера («твіст» структури), утворюючи жорстку «основу» , з якої пов'язані інші ділянки молекули
4. / -Білки l l -Спіралі та -структури чергуються по ходу ланцюга Більшість -структур (паралельні) виявляються локалізованими в центральній частині молекули, де ці структури згинаються у вигляді пропелера («твіст» структури), утворюючи жорстку «основу» , з якої пов'язані інші ділянки молекули


 Домени l l l Середній розмір домену зазвичай – 100 – 150 залишків, що відповідає глобулі з діаметром близько 2, 5 нм. е. утворення третинної структури, що відбувається на заключних етапах формування глобули
Домени l l l Середній розмір домену зазвичай – 100 – 150 залишків, що відповідає глобулі з діаметром близько 2, 5 нм. е. утворення третинної структури, що відбувається на заключних етапах формування глобули

 Третинна структура білка… l l l характеризує просторове розташування впорядкованих та аморфних ділянок у поліпептидному ланцюгу в цілому, яке досягається за рахунок взаємодії бічних радикалів і залежить від їх типу та конформації Таким чином, третинна структура описує просторове укладання всієї молекули білка, якщо вона утворена однією поліпептидною Має пряме відношення до форми молекул білка, яка може бути різною: від кулястої до ниткоподібної
Третинна структура білка… l l l характеризує просторове розташування впорядкованих та аморфних ділянок у поліпептидному ланцюгу в цілому, яке досягається за рахунок взаємодії бічних радикалів і залежить від їх типу та конформації Таким чином, третинна структура описує просторове укладання всієї молекули білка, якщо вона утворена однією поліпептидною Має пряме відношення до форми молекул білка, яка може бути різною: від кулястої до ниткоподібної
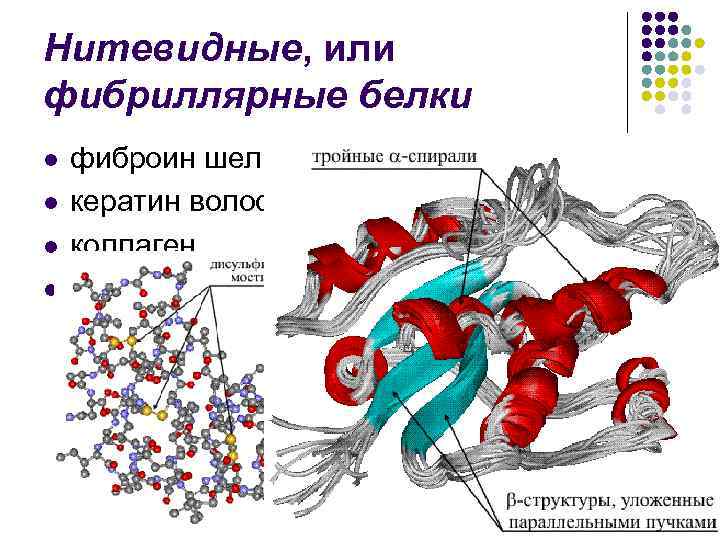 Ниткоподібні, або фібрилярні білки l l фіброїн шовку кератин волосся, рогів, копит колаген та ін.
Ниткоподібні, або фібрилярні білки l l фіброїн шовку кератин волосся, рогів, копит колаген та ін.

 Третинна структура білка l l основа функціональності білка, яка вимагає точної просторової організації великих ансамблів, побудованих з безлічі амінокислотних залишків. білка (денатурація) незмінно призводить до втрати ним можливості функціонувати
Третинна структура білка l l основа функціональності білка, яка вимагає точної просторової організації великих ансамблів, побудованих з безлічі амінокислотних залишків. білка (денатурація) незмінно призводить до втрати ним можливості функціонувати
 Третинна структура білка l l l Стабільність третинної структури залежить від системи нековалентних взаємодій усередині білкової глобули (?) Деякі білки додатково стабілізуються ковалентними – дисульфідними – зв'язками Gланцюг = RТln. К = Нцепь – Т S ланцюг Нцепь 0 (S
Третинна структура білка l l l Стабільність третинної структури залежить від системи нековалентних взаємодій усередині білкової глобули (?) Деякі білки додатково стабілізуються ковалентними – дисульфідними – зв'язками Gланцюг = RТln. К = Нцепь – Т S ланцюг Нцепь 0 (S
 Третинна структура білка l Співвідношення Нцепь і -Т Sчіп залежить від: l l числа нековалентних зв'язків, що встановлюються при згортанні молекули білка в глобулу, а від довжини поліпептидного ланцюга від температури. Зі зростанням Т |Т S| рано чи пізно перевищить | Н| та просторова структура втратить стабільність – теплова денатурація білка
Третинна структура білка l Співвідношення Нцепь і -Т Sчіп залежить від: l l числа нековалентних зв'язків, що встановлюються при згортанні молекули білка в глобулу, а від довжини поліпептидного ланцюга від температури. Зі зростанням Т |Т S| рано чи пізно перевищить | Н| та просторова структура втратить стабільність – теплова денатурація білка
 Третинна структура білка l l При згортанні глобули зменшується ентропія пептидного ланцюга, але одночасно відбувається зростання ентропії розчинника – води Це відіграє вирішальну роль у стабілізації третинної структури
Третинна структура білка l l При згортанні глобули зменшується ентропія пептидного ланцюга, але одночасно відбувається зростання ентропії розчинника – води Це відіграє вирішальну роль у стабілізації третинної структури
 Третинна структура білка l l Вода – незвичайний, високою мірою структурований розчинник (?) Полярні групи білка легко включаються до льодоподібних структур води, встановлюючи з нею водневі зв'язки Неполярні групи займають своєрідні порожнини, утворені молекулами води, пов'язаними водневими зв'язками Все це виключає з хаотичного руху значна кількість молекул води, тобто відбувається впорядкування, зменшення ентропії води
Третинна структура білка l l Вода – незвичайний, високою мірою структурований розчинник (?) Полярні групи білка легко включаються до льодоподібних структур води, встановлюючи з нею водневі зв'язки Неполярні групи займають своєрідні порожнини, утворені молекулами води, пов'язаними водневими зв'язками Все це виключає з хаотичного руху значна кількість молекул води, тобто відбувається впорядкування, зменшення ентропії води
 Третична структура білка l l l Але оскільки неполярні групи білка не можуть «відшаровуватися» від молекули, відбувається згортання глобули, при якому значна частина (не менше ½) гідрофобних груп виявляється прихованою від контакту з водою. Це тягне за собою встановлення гідрофобних контактів, ван-дер-ваальсових. сил Відбувається зниження ентропії білка Наслідком утворення глобули є руйнація значної частини водневих зв'язків та гідрофобних взаємодій бічних груп з молекулами води Зростає хаотичність руху останніх, і ентропія води зростає
Третична структура білка l l l Але оскільки неполярні групи білка не можуть «відшаровуватися» від молекули, відбувається згортання глобули, при якому значна частина (не менше ½) гідрофобних груп виявляється прихованою від контакту з водою. Це тягне за собою встановлення гідрофобних контактів, ван-дер-ваальсових. сил Відбувається зниження ентропії білка Наслідком утворення глобули є руйнація значної частини водневих зв'язків та гідрофобних взаємодій бічних груп з молекулами води Зростає хаотичність руху останніх, і ентропія води зростає

 Четвертична структура білка… l l l Четвертинну структуру мають олігомерні білки, молекули яких складаються з двох і більше поліпептидних ланцюгів, пов'язаних нековалентно. ансамбль глобул) виступає як єдина молекула, в той же час кожна з глобулів, що об'єдналися, зберігає значну автономність.
Четвертична структура білка… l l l Четвертинну структуру мають олігомерні білки, молекули яких складаються з двох і більше поліпептидних ланцюгів, пов'язаних нековалентно. ансамбль глобул) виступає як єдина молекула, в той же час кожна з глобулів, що об'єдналися, зберігає значну автономність.

 Четвертична структура білка l l Субодиниці, що утворюють четвертинну структуру білка, можуть бути абсолютно різними за будовою та функціями – гетеромерні білки Це дозволяє об'єднати в одній структурі кілька взаємозалежних функцій, створити поліфункціональну молекулу l l Приклад. Протеїнкіназа: С-субодиниця відповідає за ферментативну активність, а R-субодиниця – регуляторна У гомомірних білках субодиниці однакові
Четвертична структура білка l l Субодиниці, що утворюють четвертинну структуру білка, можуть бути абсолютно різними за будовою та функціями – гетеромерні білки Це дозволяє об'єднати в одній структурі кілька взаємозалежних функцій, створити поліфункціональну молекулу l l Приклад. Протеїнкіназа: С-субодиниця відповідає за ферментативну активність, а R-субодиниця – регуляторна У гомомірних білках субодиниці однакові
 Четвертична структура білка l Міжсубодиничні контакти – дуже розвинена система нековалентних взаємодій Четвертична структура менш міцна, ніж третинна, тому що менше внесок гідрофобних контактів
Четвертична структура білка l Міжсубодиничні контакти – дуже розвинена система нековалентних взаємодій Четвертична структура менш міцна, ніж третинна, тому що менше внесок гідрофобних контактів

 Функції четвертинної структури 1. Об'єднання кількох взаємопов'язаних функцій в єдиній структурі 2. Архітектурна функція 1. Формування просторових утворень дуже складної конфігурації, що забезпечують специфічні функціональні можливості білка (феритин) 2. Сумарне проведення послідовних реакцій ферментів 3. Формування функціональних
Функції четвертинної структури 1. Об'єднання кількох взаємопов'язаних функцій в єдиній структурі 2. Архітектурна функція 1. Формування просторових утворень дуже складної конфігурації, що забезпечують специфічні функціональні можливості білка (феритин) 2. Сумарне проведення послідовних реакцій ферментів 3. Формування функціональних
 Функції четвертинної структури 3. Забезпечення множинних взаємодій білка з протяжними структурами ДНК-зв'язуючі білки – димери (?) 4. Регуляторна функція. Передача ефекту (порушення третинної структури при взаємодії з субстратом) передається від однієї субодиниці до іншої, що призводить до перебудови всієї четвертинної структури
Функції четвертинної структури 3. Забезпечення множинних взаємодій білка з протяжними структурами ДНК-зв'язуючі білки – димери (?) 4. Регуляторна функція. Передача ефекту (порушення третинної структури при взаємодії з субстратом) передається від однієї субодиниці до іншої, що призводить до перебудови всієї четвертинної структури


 1. Кожен індивідуальний білок характеризується унікальною структурою, що забезпечує унікальність його функцій 2. Тому з'ясування структури різноманітних білків може бути ключем до пізнання природи живих систем і, відповідно, сутності
1. Кожен індивідуальний білок характеризується унікальною структурою, що забезпечує унікальність його функцій 2. Тому з'ясування структури різноманітних білків може бути ключем до пізнання природи живих систем і, відповідно, сутності
 Література Березов Т. Т., Коровкін Б. Ф. Біологічна хімія. - М.: Медицина, 1983 Мецлер Д. Біохімія. Хімічні реакції у живій клітині. У 3-х т. - Т. т. 1, 2. - М.: Світ, 1980 Овчинніков Ю. А. Біоорганічна хімія. - М.: Просвітництво, 1987 Основи біохімії / За ред. А. А. Анісімова. - М.: Вища школа, 1986 Рис Е., Стренберг М. Від клітин до атомів. Ілюстроване введення молекулярну біологію. - М.: Світ, 1988 Степанов В. М. Молекулярна біологія. Структура та функції білків. - М.: Вища школа, 1996 Пилипович Ю. Б. Основи біохімії. - М.: Вища школа, 1993
Література Березов Т. Т., Коровкін Б. Ф. Біологічна хімія. - М.: Медицина, 1983 Мецлер Д. Біохімія. Хімічні реакції у живій клітині. У 3-х т. - Т. т. 1, 2. - М.: Світ, 1980 Овчинніков Ю. А. Біоорганічна хімія. - М.: Просвітництво, 1987 Основи біохімії / За ред. А. А. Анісімова. - М.: Вища школа, 1986 Рис Е., Стренберг М. Від клітин до атомів. Ілюстроване введення молекулярну біологію. - М.: Світ, 1988 Степанов В. М. Молекулярна біологія. Структура та функції білків. - М.: Вища школа, 1996 Пилипович Ю. Б. Основи біохімії. - М.: Вища школа, 1993


