Struktura jednog proteina određena je skupinom gena. Proteini: primarna struktura proteina, shema sastava tripeptida
Bilki- Visokomolekularni organski spojevi koji nastaju iz viška α-aminokiselina.
U skladište proteina uključuju ugljen, vodu, dušik, kisen, sirku. Neki proteini tvore komplekse s drugim molekulama, poput fosfora, željeza, cinka i bakra.
Proteini imaju visoku molekulsku težinu: albumin jaja - 36 000, hemoglobin - 152 000, miozin - 500 000. Za usporedbu: molekulska težina alkohola - 46, ušna kiselina - 60, benzen - 78.
Skladište aminokiselina proteina
Bilki- Neperiodični polimeri, kao što su monomeri α-aminokiseline. Postoji 20 vrsta α-aminokiselina koje se nazivaju proteinski monomeri, iako ih je preko 170 identificirano u stanicama i tkivima.
Važno je napomenuti da se aminokiseline mogu sintetizirati u ljudskom tijelu i drugim bićima, a dijele se na: zamjenske aminokiseline- Može se sintetizirati; esencijalne aminokiseline- Ne mogu sintetizirati. Esencijalne aminokiseline nalaze se u tijelu izravno iz tijela. Biljke sintetiziraju sve vrste aminokiselina.
Pohranjeno u skladištu aminokiselina, proteini su u punom jeku- Raspršite cijeli set aminokiselina; inferioran- Sve aminokiseline su dnevno na zalihama. Budući da se proteini sastoje od aminokiselina, tzv žao mi je. Kada proteini aminokiselinama dodaju još jednu komponentu koja nije aminokiselina (prostetska skupina), nazivaju se sklopivi. Protetičku skupinu mogu predstavljati metali (metaloproteini), ugljikohidrati (glikoproteini), lipidi (lipoproteini), nukleinske kiseline (nukleoproteini).
Brkovi aminokiseline se osvetiti: 1) karboksilna skupina (-COOH); 2) amino skupina (-NH2); 3) radikal ili R-skupina (reshta molekula). Budov radikal varira u različitim vrstama aminokiselina. Ovisno o broju amino skupina i karboksilnih skupina koje ulaze u skladište aminokiselina, dijele se na: neutralne aminokiseline Postoji jedna karboksilna skupina i jedna amino skupina; bazične aminokiseline Postoji više od jedne amino skupine; kisele aminokiselineŠto je više od jedne karboksilne skupine?
Aminokiseline amfoterni poluvodiči Dakle, u hrani smrad mogu uzrokovati i kiseline i baze. U vodenim varijantama aminokiseline se pojavljuju u različitim ionskim oblicima.
Peptidna veza
Peptidi- Organski spojevi koji nastaju od viška aminokiselina spojenih peptidnom vezom.
Oslobađanje peptida slijedi reakciju kondenzacije aminokiselina. Kada amino skupina jedne aminokiseline stupa u interakciju s karboksilnom skupinom druge, između njih nastaje kovalentna veza dušik-ugljikohidrat, tzv. peptid. Važno je odvojiti količinu viška aminokiselina koja odlazi u skladište peptida dipeptidi, tripeptidi, tetrapeptidi itd. Stvaranje peptidne veze može se ponoviti na mnogo načina. Iznesite ovo na vidjelo polipeptidi. Na jednom kraju peptida nalazi se slobodna amino skupina (nazvana N-kraj), a na drugom kraju je slobodna karboksilna skupina (nazvana C-kraj).

Jednostavna organizacija proteinskih molekula
Specifične funkcije proteina ovise o prostornoj konfiguraciji njihovih molekula, osim toga, stanice energetski i nevidljivo režu proteine u otvorenom obliku, koji izgleda poput koplja, pa se polipeptidna koplja uočava stvaranje i bubrenje trivijalne strukture i konformacije. Pogledajte 4 regije prostrana organizacija proteina.
Primarna struktura proteina- Slijed otapanja viška aminokiselina iz polipeptidne lancete da bi se formirala proteinska molekula. Veza između aminokiselina je peptidna.

Budući da se proteinska molekula sastoji od samo 10 viška aminokiselina, tada su teoretski moguće varijante proteinskih molekula koje variraju u redoslijedu dodavanja aminokiselina 10 20. Uz 20 aminokiselina, možete ih kombinirati u još više različitih kombinacija. Kod ljudi je identificirano oko deset tisuća različitih proteina koji se razlikuju od jedne do druge vrste, kao i od proteina drugih organizama.
Sama primarna struktura proteinske molekule ukazuje na snagu proteinske molekule i njenu prostranu konfiguraciju. Zamjena jedne aminokiseline drugom u polipeptidnom lancu dovodi do promjene u moći i funkcijama proteina. Na primjer, zamjena glutaminske aminokiseline u β-podjedinici hemoglobina s valinom dovodi do činjenice da molekula hemoglobina kao cjelina ne može izgubiti svoju glavnu funkciju - transport kiseline; U takvim slučajevima ljudi razvijaju bolest - anemiju srpastih stanica.
Sekundarna struktura- Larinks polipeptidne lancete je posložen u spiralu (izgleda kao proširena opruga). Zavojnice spirale tvore vodene veze koje se isprepliću između karboksilnih skupina i amino skupina. U stvaranju vodnih veza sudjeluju gotovo sve CO- i NH-skupine. Smrad je slabiji od peptidnog, ali je pri ponavljanju bogat plinovima, što ovim promjenama daje postojanost i oštrinu. Na razini sekundarne strukture nalaze se proteini: fibroin (šav, svila), keratin (kosa, nokti), kolagen (tetiva).



Tretinska struktura- Djevojka polu-petd Lancyugiv na Globuli, viniki veneolikenny od vinikenni devedesetih Zv'yazkiv (vodnevikh, disulfídni) je nanošenje girdrophic međusobnog mira radikalima aminokiselina. Glavnu ulogu osvijetljene tercijarne strukture imaju hidrofilno-hidrofobne interakcije. U vodenim molekulama, hidrofobni radikali nastoje konvergirati u vodi, grupirajući se u sredini globule, dok hidrofilni radikali, kao rezultat hidratacije (interakcija s dipolima vode), nastoje se taložiti na površini molekule. U nekim je proteinima tercijarna struktura stabilizirana disulfidnim kovalentnim vezama koje nastaju između ugljikovih atoma dva suvišna cisteina. Na razini tercijarne strukture nalaze se enzimi, antitijela i određeni hormoni.

Kvartarna struktura a kod savijanja proteina, molekule koje se formiraju u dvije ili više kuglica. Podjedinice se kombiniraju u molekuli kroz ionske, hidrofobne i elektrostatske interakcije. Ponekad, kada se formira kvaternarna struktura, pojavljuju se disulfidne veze između podjedinica. Najveći protein koji ima četvrtinsku strukturu je hemoglobin. Sadrži dvije α-podjedinice (141 aminokiselina u višku) i dvije β-podjedinice (146 aminokiselina u višku). Molekula hema povezana je s podjedinicom kože, koja je odgovorna za oslobađanje.
Ako iz bilo kojeg razloga konformacija proteina odstupi od normalne, protein ne može zadržati svoje funkcije. Na primjer, uzrok "kravlje priče" (spongiformna encefalopatija) je abnormalna konformacija priona - površinskih proteina živčanih stanica.
Snaga vjeverica
Skladištenje aminokiselina, struktura proteinske molekule znači vlast. Proteini dobivaju svoju bazičnu kiselost, koja je određena radikalima aminokiselina: što protein ima kiselije aminokiseline, to je izraženiji izraz njegove kiselosti. Vrijednost davanja i dolaska H+ znači puferska snaga proteina; Jedan od najvažnijih pufera je hemoglobin u crvenim krvnim stanicama, koji održava pH krvi na konstantnoj razini. - odvojene bjelančevine (fibrinogen) i neodvojene bjelančevine koje doprinose mehaničkim funkcijama (fibrin, keratin, kolagen). Postoje aktivni proteini u kemijskom obliku (enzimi), a postoje i oni kemijski neaktivni, otporni na prodor različitih umova iz vanjskog svijeta i krajnje nestabilni.
Vanjski čimbenici (grijanje, izloženost ultraljubičastom zračenju, važni metali i njihove soli, promjene pH vrijednosti, zračenje, zagađenje vode)
može uzrokovati oštećenje strukturne organizacije proteinske molekule. Proces gubitka trivijalne konformacije, dominantne konformacije proteinske molekule, naziva se denaturacija. Razlog denaturacije je pucanje ligamenata koji stabiliziraju strukturu proteina. U bubregu pucaju najslabiji ligamenti, a kad su pameti jače, pucaju i jači. Zatim se razvijaju kvartarne, zatim tercijarne i sekundarne strukture. Promjena prostorne konfiguracije dovodi do promjene snage proteina i, posljedično, onemogućuje gubitak kontrole nad svojim biološkim funkcijama. Budući da denaturacija nije popraćena razaranjem primarne strukture, može vukodlak, nakon što dođe do samoobnavljanja konformacije proteina snage. Takvu denaturaciju prepoznaju, na primjer, membranski receptorski proteini. Proces obnove strukture proteina nakon denaturacije naziva se renaturacija. Ako je nemoguće obnoviti prostornu konfiguraciju proteina, tada se zove denaturacija neopozivo.
Funkcije proteina
| Funkcija | Molimo navedite objašnjenje |
|---|---|
| Budivelna | Proteini sudjeluju u formiranju staničnih i postkliničkih struktura: ulaze u stanične membrane (lipoproteini, glikoproteini), kosu (keratin), tetive (kolagen) itd. |
| Transportna | Bjelančevina krvi hemoglobin dodaje kiselost i prenosi je iz krvi u sva tkiva i organe, a iz njih prenosi ugljični dioksid; Skladištenje staničnih membrana uključuje posebne proteine koji osiguravaju aktivan i strogo selektivan prijenos određenih tvari i iona iz stanice u vanjsku jezgru i natrag. |
| Regulatorni | Proteinski hormoni sudjeluju u regulaciji procesa metabolizma govora. Na primjer, hormon inzulin regulira razinu glukoze u krvi, pospješuje sintezu glikogena i povećava redukciju masti iz ugljikohidrata. |
| Zahisna | Prodiranjem stranih proteina i mikroorganizama (antigena) u organizam stvaraju se posebni proteini - antitijela, koja ih vežu i ljušte. Fibrin, koji se stvara s fibrinogenom, smanjuje krvarenje. |
| Dviguna | Kratkotrajni proteini aktin i miozin osiguravaju skraćivanje ulkusa kod životinja bogatih stanicama. |
| Signalna | Površinska membrana stanice sadrži proteinske molekule koje mijenjaju svoju tercijarnu strukturu kao odgovor na čimbenike vanjskog medija, čime primaju signale iz vanjskog medija i prenose naredbe stanici. |
| Stvaranje zaliha | U tijelu životinja, bjelančevine se, u pravilu, ne pohranjuju, krivci: albumin jaja, kazein mlijeka. No, bjelančevine u tijelu mogu se opskrbiti rezervom određene količine govora, npr. pri raspadu hemoglobina tekućina se ne izlučuje iz tijela, već se pohranjuje, stvarajući kompleks s bjelančevinom feritinom. |
| energično | Pri razgradnji 1 g bjelančevina do krajnjih produkata vidljivo je 176 kJ. U početku se proteini razgrađuju na aminokiseline, a zatim na krajnje proizvode - vodu, ugljični dioksid i amonijak. Međutim, kao izvor energije, proteini se apsorbiraju samo ako se konzumiraju drugi izvori (ugljikohidrati i masti). |
| Katalitički | Jedna od najvažnijih funkcija proteina. Opskrbljen proteinima - enzimima koji ubrzavaju biokemijske reakcije koje se odvijaju u stanicama. Na primjer, ribuloza bifosfat karboksilaza katalizira fiksaciju 2 tijekom fotosinteze. |
Fermenti
Fermenti, ili enzima, - Posebna klasa proteina koji su biološki katalizatori. Zbog enzima, biokemijske reakcije odvijaju se velikom fluidnošću. Brzina enzimskih reakcija u desecima tisuća puta (i u milijunima puta) posljedica je brzine reakcija koje se odvijaju uz sudjelovanje anorganskih katalizatora. Rechovina, što je ono što enzim daje svoj učinak, zove se supstrat.
Enzimi - globularni proteini, osobitosti Budovi Enzime možemo podijeliti u dvije skupine: jednostavne i složene. Žao mi je ferment s najjednostavnijim proteinima, dakle. nastaju prvenstveno od aminokiselina. Preklopni enzimiê presavijanje bijelih, zatim. Prije njihovog skladišta, osim proteinskog dijela, postoji skupina neproteinske prirode. kofaktor. U nekim enzimima vitamini djeluju kao kofaktori. Molekula enzima ima poseban dio koji se naziva aktivni centar. Aktivni centar- Mali dio enzima (od tri do dvanaest viška aminokiselina), koji veže supstrat ili supstrate u kompleks enzim-supstrat. Nakon završetka reakcije, kompleks enzim-supstrat se razgrađuje na enzim i proizvod(e) reakcije. Stvarni enzimi su aktivni (aktivna krema) alosterički centri- Dijelovi za koje se koriste regulatori likvidnosti i enzimi ( alosterički enzimi).

Reakcije enzimske katalize karakteriziraju: 1) visoka učinkovitost, 2) stroga selektivnost i izravnost djelovanja, 3) specifičnost supstrata, 4) fina i precizna regulacija. Specifičnost supstrata i reakcije reakcija enzimske katalize objašnjena je hipotezama E. Fishera (1890.) i D. Koshlanda (1959.).
E. Fisher (hipoteza ključ-brava) Pod pretpostavkom da su široke konfiguracije aktivnog centra enzima i supstrata potpuno iste. Supstrat je isti kao "ključ", enzim je "brava".
D. Koshland (hipoteza rukavica) Pod pretpostavkom da se sličnost strukture između supstrata i aktivnog centra enzima stvara samo u trenutku njihove interakcije jednog po jednog. Ova se hipoteza također naziva hipoteza induciranih vrsta.
Fluidnost enzimskih reakcija ovisi o: 1) temperaturi; 2) koncentracija enzima; 3) koncentracija supstrata; 4) pH. Valja napomenuti da su fragmenti enzima proteini, tada je njihova aktivnost najveća u fiziološki normalnim umovima.
Većina enzima može se obraditi samo na temperaturama u rasponu od 0 do 40 °C. U tim se područjima fluidnost reakcije povećava otprilike 2 puta kada temperatura kože poraste na 10 °C. Na temperaturama iznad 40 ° C, protein prolazi kroz denaturaciju i aktivnost enzima se smanjuje. Na temperaturama blizu točke smrzavanja enzimi se inaktiviraju.
S povećanom količinom supstrata fluidnost enzimske reakcije raste sve dok se broj molekula u supstratu ne izjednači s brojem molekula u enzimu. S daljnjim povećanjem količine supstrata, likvidnost se ne povećava, sve dok se aktivni centri enzima zasiti. Veće koncentracije enzima dovode do povećane katalitičke aktivnosti, jer u jednom satu reakcije više molekula biva izloženo supstratu.

Za kožni enzim postoji optimalna pH vrijednost pri kojoj on pokazuje maksimalnu aktivnost (pepsin – 2,0, sluzna amilaza – 6,8, potkožna lipaza – 9,0). Pri višim ili nižim pH vrijednostima aktivnost enzima opada. U slučaju velikih promjena u pH, enzim denaturira.
Fluidnost alosteričnih enzima regulirana je tvarima koje dopiru do alosteričkih centara. Budući da te riječi ubrzavaju reakciju, nazivaju se smradovima aktivatori, yakscho to galm inhibitori.
Podjela enzima
Ovisno o vrsti kataliziranih kemijskih reakcija, enzimi se dijele u 6 klasa:
- oksireduktaza(prijenos atoma iz vode, kiseline ili elektrona s jedne tvari na drugu - dehidrogenaza),
- transferasi(prijenos metilne, acilne, fosfatne ili amino skupine s jedne molekule na drugu - transaminaza),
- hidrolaze(reakcije hidrolize, u kojima se iz supstrata sintetiziraju dva produkta - amilaza, lipaza),
- liazi(nehidrolitička adicija na supstrat ili odvajanje od nove skupine atoma, pri čemu dolazi do kidanja veza C-C, C-N, C-O, C-S dekarboksilaze),
- izomeraza(Intramolekularna perebudova - izomeraza),
- ligazi(Nastanak dviju molekula kao rezultat stvaranja veza C-C, C-N, C-O, C-S - sintetaza).
Klasificirajte svoje crteže u potklase i potklase. U međunarodnoj klasifikaciji, koja postoji, kožni enzim ima jedinstveni kod koji se sastoji od četiri broja odvojena točkama. Prvi broj je klasa, drugi je podklasa, treći je podklasa, četvrti je serijski broj enzima u toj podklasi, npr. kod arginaze je 3.5.3.1.
Ići predavanja br.2"Koje su funkcije ugljikohidrata i lipida"
Ići predavanja br.4“Funkcije nukleinskih kiselina ATP”
Biosinteza proteina.
1. Struktura jednog proteina je određena:
1) skupina gena 2) jedan genom
3) jedna molekula DNA 4) ukupnost gena u organizmu
2. Gen kodira informacije o slijedu monomera u molekuli:
1) tRNA 2) AA 3) glikogen 4) DNA
3. Trojke se nazivaju antikodoni:
1) DNA 2) t-RNA 3) i-RNA 4) r-RNA
4. Plastična izmjena se značajno razvija reakcijom:
1) dezintegracija organskih govora 2) dezintegracija anorganskih govora
3) sinteza organskih tvari 4) sinteza anorganskih tvari
5. Sinteza proteina u prokariotskim stanicama odvija se:
1) na ribosomima u jezgri 2) na ribosomima u citoplazmi 3) na mjestu stanice
4) na vanjskoj površini citoplazmatske membrane
6. Proces emitiranja je pokrenut:
1) u citoplazmi 2) u jezgri 3) u mitohondrijima
4) na membranama kratke endoplazmatske membrane
7. Na membranama granularne endoplazmatske membrane dolazi do sinteze:
1) ATP; 2) u ugljikohidratima; 3) lipidi; 4) bjelančevine.
8. Jedan triplet kodova:
1. jedan AK 2 jedan znak tijela 3. udlaga AK
9. Sinteza proteina je trenutno završena
1. prepoznavanje kodona antikodonom 2. pojava “razdjelne oznake” na ribosomu
3. Prijenos i-RNA do ribosoma
10. Proces kojim se informacije čitaju s molekula DNA.
1. prijevod 2. transkripcija 3. transformacija
11. Snaga proteina je određena...
1. sekundarna struktura proteina 2. primarna struktura proteina
3. tercijarna struktura proteina
12. Proces kojim se antikodon prepoznaje kao kodon na i-RNA
13. Faze biosinteze proteina.
1.transkripcija, prijevod 2.transformacija, prijevod
3.transorganizacija, transkripcija
14. Antikodon t-RNA sastoji se od UCG nukleotida. Koji DNA triplet je komplementaran youmu?
1.UUG 2. TTC 3. TCG
15. Broj t-RNA koje sudjeluju u translaciji je tradicionalan:
1. Kodoni i-RNA koji šifriraju aminokiseline 2. molekule i-RNA
3 Geni koji su uključeni prije molekule DNA 4. Proteini koji se sintetiziraju na ribosomima
16. Odredite slijed nukleotida i-RNA tijekom transkripcije s jednog od lanaca DNA: A-G-T-C-G
1) U 2) G 3) C 4) A 5) C
17. Tijekom replikacije molekule DNA nastaje:
1) nit koja se raspala na fragmente molekula kćeri
2) molekula koja se sastoji od dva nova DNA lanca
3) molekula, čiju polovicu čine iRNA niti
4) molekula kćer koja nastaje od jednog starog i jednog novog lanca DNK
18. Predložak za sintezu molekule iRNA tijekom transkripcije je:
1) cijela molekula DNA 2) površina jedne od molekula DNA
3) zaplet jednog od Lancsovih DNK
4) kod nekih vrsta jedna molekula DNK, kod drugih – cijela molekula DNK.
19. Proces samosubdukcije molekule DNA.
1. replikacija 2. popravak
3. reinkarnacija
20. Tijekom biosinteze proteina u stanicama, ATP energija:
1) potrošiti 2) opskrbiti se
3) nije potrošeno i nije viđeno
21. U somatskim stanicama organizma bogatog stanicama:
1) drugačiji skup gena i proteina 2) novi skup gena i proteina
3) novi set gena, ili drugačiji skup proteina
4) novi set proteina, ili drugačiji skup gena
22.. Jedan triplet DNK nosi informacije o:
1) slijed aminokiselina u proteinskoj molekuli
2) znakovi tijela 3) aminokiseline u sintetiziranoj proteinskoj molekuli
4) skladištenje molekule RNA
23. Koji od procesa nije prisutan u stanicama niti jedne funkcije:
1) sinteza proteina 2) metabolizam proteina 3) mitoza 4) mejoza
24. Koncept "transkripcije" odnosi se na proces:
1) subdukcija DNA 2) sinteza i-RNA na DNA
3) prijelaz i-RNA u ribosome 4) stvaranje proteinskih molekula na polisomima
25. Dio molekule DNA koji nosi informaciju o jednoj proteinskoj molekuli:
1) gen 2) fenotip 3) genom 4) genotip
26. Transkripcija u eukariota događa se u:
1) citoplazma 2) endoplazmatska membrana 3) lizosomi 4) jezgre
27. Sinteza proteina se dobiva iz:
1) granularni endoplazmatski retikulum
2) glatki endoplazmatski retikulum 3) jezgra 4) lizosomi
28. Jedna aminokiselina je kodirana:
1) nekoliko nukleotida 2) dva nukleotida
3) jedan nukleotid 4) tri nukleotida
29. Triplet ATC nukleotida u molekuli DNA ima kodon sličan molekuli i-RNA:
1) TAG 2) UAG 3) UTC 4) TsAU
30. Razdjelni znakovigenetski kod:
1. kodiraju proteine pjesme 2. pokreću sintezu proteina
3. pospješuju sintezu proteina
31. Proces samoprepisivanja molekule DNA.
1.replikacija 2.popravak 3.reankornacija
32. Funkcija i-RNA u procesu biosinteze.
1.spremanje informacija o zbijanju 2.transport AK do ribosoma
3. opskrba ribosomima informacijama
33. Proces kada t-RNA dovodi aminokiseline do ribosoma.
1.prijepis 2.prijevod 3.transformacija
34. Ribosomi, koji sintetiziraju istu proteinsku molekulu.
1.kromosom 2.polisom 3.megakromosom
35. Proces kojim aminokiseline stabiliziraju proteinsku molekulu.
1.prijepis 2.prijevod 3.transformacija
36. Prije reakcije matrične sinteze donosimo...
1. replikacija DNK 2. transkripcija, translacija 3. dvije vrste su točne
37. Jedan triplet DNK nosi informacije o:
1. Redoslijed aminokiselina u proteinskoj molekuli
2. Mjesto pjevanja AK u proteinskoj lanceti
3. Znak određenog organizma
4. Aminokiseline koje su uključene u lancet proteina
38. Genya je kodirao podatke o:
1) struktura bjelančevina, masti i ugljikohidrata 2) primarna struktura bjelančevina
3) slijed nukleotida u DNA
4) slijed aminokiselina u 2 ili više proteinskih molekula
39. Sinteza mRNA počinje:
1) cijepanje DNA u dva lanca 2) interakcija s enzimom RNA polimerazom i genom
3) subgeneracija gena 4) raspad gena na nukleotide
40. Transkripcija je osigurana:
1) u jezgri 2) na ribosomima 3) u citoplazmi 4) na glatkim EPS kanalima
41. Sinteza proteina ne događa se na ribosomima u:
1) zbudnik tuberkulozni 2) bjoli 3) muhara 4) bakteriofag
42. Tijekom translacije, matrica za sastavljanje polipeptidnog proteina je:
1) jedan od DNA lanaca 2) jedan od DNA molekula
3) molekula iRNA 4) u nekima čini jedan od lanaca DNA, u drugima – molekula iRNA
Primarna struktura proteina je linearno polipeptidno koplje s aminokiselinama povezanim peptidnim vezama. Primarna struktura je najjednostavnija razina strukturne organizacije proteinske molekule. Visoku stabilnost osiguravaju kovalentne peptidne veze između α-amino skupine jedne aminokiseline i α-karboksilne skupine druge aminokiseline.
Ako u peptidnoj vezi sudjeluje amino skupina prolina ili hidroksiprolina, onda izgleda drugačije

Kada se u bubregu stvaraju peptidne veze, karboksilna skupina jedne aminokiseline se aktivira, a zatim se spaja s drugom amino skupinom. Također je moguće provesti laboratorijsku sintezu polipeptida.
Peptidna veza je fragment polipeptidnog koplja koji se ponavlja. Postoji nekoliko posebnosti koje se odnose ne samo na oblik primarne strukture, već i na najvažniju razinu organizacije polipeptidne lancete:


· Koplanarnost - svi atomi koji su uključeni u peptidnu skupinu nalaze se u istoj ravnini;
· Valjanost je prisutna u dva rezonantna oblika (keto i enolni oblici);
· transpozicija intercesora u stopostotnoj C-N vezi;
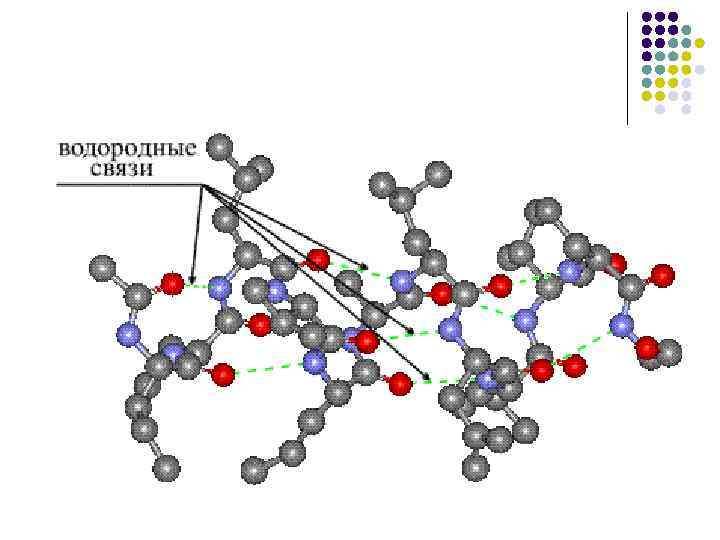
· Prisutnost formiranja vodenih ligamenata, pri čemu koža s peptidnim skupinama može stvoriti dva vodena ligamenta s drugim skupinama, uključujući i peptidne.
Peptidne skupine su povezane s amino skupinom prolina ili hidroksiprolina. Smrad zgrade stvara samo jedan vodenasti zvuk (divna stvar). To je naznačeno na oblikovanoj sekundarnoj strukturi proteina. Polipeptidni spoj, koji sadrži prolin ili hidroksiprolin, lako se uništava i ne može se ukloniti, kao i uvijek, drugim vodenim vezama.
shema formulacije tripeptida:

Razine prostorne organizacije proteina: sekundarna struktura proteina: koncepti α-heliksa i β-nabrane lopte. Tercijarna struktura proteina: pojmovi o nativnom proteinu i denaturaciji proteina. Kvartarna struktura proteina je slična hemoglobinu.
Sekundarna struktura proteina. Pod sekundarnom strukturom proteina podrazumijevamo način na koji je polipeptidno koplje raspoređeno u uređenu strukturu. Prema konfiguraciji vidljivi su sljedeći elementi sekundarne strukture: α -spirala β - Lopta za dijelove skladišta.
Budovi model α-spirale, koji štiti svu snagu peptidne veze, razbili su L. Pauling i R. Corey (1949. - 1951.).
Bebi 3, A prikazan dijagram α -spirale, koji daje informacije o glavnim parametrima polipeptidne lancete koja gori α - spiralu na način da su zavoji spirale pravilni, tako da konfiguracija spirale ima vijčanu simetriju (sl. 3, b). na kožni turn α -spirale imaju 3,6 aminokiselina viška. Stanite između zavoja ili rub spirale postaje 0,54 nm, gdje sljedeći zavoj doseže 26 °. Oblikovanje i podrezivanje α -spiralna konfiguracija nastaje građom vodenih ligamenata koji se stvaraju između peptidnih skupina kože n-idi tamo ( P+ 3) višak aminokiselina. Iako je energija vodenih ligamenata mala, njihova je količina velika da proizvede značajan energetski učinak, uslijed čega α - Spiralna konfiguracija za kompletiranje postolja. Biološki radikali viškova aminokiselina ne sudjeluju u tvari α -spiralna konfiguracija, dakle svi viškovi aminokiselina u α -Spirale su jednake.
Prirodni proteini su manje desnokretni α - Spirale.
β-savijena lopta- Još jedan element sekundarne strukture. Na stranici administracije α -spirale β -Presavijena lopta ima linearni oblik, a ne strizhnevuyu oblik (slika 4). Linearna struktura određena je podrijetlom vodenih veza između peptidnih skupina koje se nalaze na različitim dijelovima polipeptidnog koplja. Čini se da su ove plohe blizu površine vodene veze između - C = O i HN - skupina (0,272 nm).

Mali 4. Shematski prikaz β
- skladišna česta lopta (strelice pokazuju
 o polipeptidnoj lanceti)
o polipeptidnoj lanceti)
Mali 3. Shema ( A) taj model ( b) α -spirale
Sekundarna struktura proteina identificirana je kao primarna struktura. Višak aminokiselina u različitim količinama stvara se prije nego što se formiraju vodeni ligamenti, koji se zatim ulijeva u smjesu. α -spirale odn β -Sharu. Aminokiseline topljive u spirali uključuju alanin, glutaminsku kiselinu, glutamin, leucin, lizin, metionin i histidin. Ako se proteinski fragment formira prvenstveno od viška aminokiselina, tada će se on formirati u ovoj fazi α -spirala. Valin, izoleucin, treonin, tirozin i fenilalanin pružaju rješenje β - Kuglice polipeptidne lancete. Na plohama polipeptidnog koplja pojavljuju se neuređene strukture, gdje su koncentrirani ostaci aminokiselina kao što su glicin, serin, asparaginska kiselina, asparagin, prolin.
U bogatim proteinima, odmah α -spirale, to β -Shari. Dio spiralne konfiguracije je različit u različitim proteinima. Dakle, mesni protein paramiozin je praktički 100% spiraliziran; visok udio spiralne konfiguracije u mioglobinu i hemoglobinu (75%). Međutim, u tripsinu i ribonukleazi značajan dio polipeptida lanjuga stane u šaruvat β -strukture. Proteini potpornih tkiva - keratin (proteini kose), kolagen (proteini kože i tetiva) - bubre β -Konfiguracija polipeptidnih koplja.
Tercijarna struktura proteina. Tercijarna struktura proteina način je polaganja polipeptidnog koplja u prostoru. Kako bi protein svojoj funkcionalnoj snazi pridodao snagu, polipeptidna lanceta je dužna pojedinačno sagorjeti u prostoru, tvoreći funkcionalno aktivnu strukturu. Ova struktura se zove domaći. Bez obzira na veliki broj prostornih struktura koje su teoretski moguće za okolnu polipeptidnu lancetu, laringealni protein treba dovesti do uspostavljanja jedinstvene nativne konfiguracije.
Stabilizirati tercijarnu strukturu proteina interakcijama koje se javljaju između bioloških radikala aminokiselinskih viška različitih dijelova polipeptidnog koplja. Ove interakcije mogu se podijeliti na prednosti i slabosti.
Snažne interakcije uključuju kovalentne veze između atoma sumpora viška cisteina, koji se nalaze na različitim dijelovima polipeptidnog koplja. Inače se takve veze nazivaju disulfidnim mostovima; Stvaranje disulfidnog mosta može se prikazati na sljedeći način:
Osim kovalentnih veza, na tercijarnu strukturu proteinske molekule utječu slabe interakcije koje se po svojoj prirodi dijele na polarne i nepolarne.
Polarne interakcije uključuju veze iona i vode. Ionske interakcije nastaju u kontaktu između pozitivno nabijenih skupina otpadnih radikala lizina, arginina, histidina i negativno nabijenih COOH skupina asparaginske i glutaminske kiseline. Vodene veze javljaju se između funkcionalnih skupina bioloških radikala viškova aminokiselina.
Nepolarne ili van der Waalsove interakcije između ugljikohidratnih radikala aminokiselinskih ostataka sprječavaju stvaranje hidrofobna jezgra (Masne mrlje) u sredini proteinske globule, jer Ugljikohidratni radikali će nestati u kontaktu s vodom. Što više nepolarnih aminokiselina protein sadrži, veća je uloga van der Waalsovih veza u njegovoj oblikovanoj tercijarnoj strukturi.
Brojčane veze između proteinskih radikala viška aminokiselina ukazuju na prostornu konfiguraciju proteinske molekule (slika 5).

Mali 5. Vrste ligamenata koji podupiru tercijarnu strukturu proteina:
A- smjesa disulfida; b - Ionska veza; c, d - priključci za vodu;
d - van der Waalsove obveznice
Tercijarna struktura novouzetog proteina je jedinstvena, kao što je jedinstvena i primarna struktura. Samo pravilan prostor za polaganje proteina će ga održati aktivnim. Masivno uništenje treće strukture dovodi do promjene razine proteina i gubitka biološke aktivnosti.
Kvartarna struktura proteina. Proteini s molekularnom težinom većom od 100 kDa 1 obično se sastoje od nekoliko polipeptidnih koplja male molekulske mase. Struktura koja se sastoji od velikog broja polipeptidnih koplja, koji zauzimaju strogo fiksiran položaj, naizgled jedan nasuprot drugog, zbog čega protein ima različitu aktivnost, naziva se kvaternarnom strukturom proteina. Protein koji ima četvrtinsku strukturu naziva se epimolekula ili drugo multimer , a skladišta njegovih polipeptidnih lanceta jasno su podjedinice ili drugo protomiri . Karakteristična snaga proteina s kvartarnom strukturom su oni čija podjedinica nema biološku aktivnost.
Stabilizacija kvaternarne strukture proteina postiže se interakcijom polarnih interakcija između radikala aminokiselina lokaliziranih na površini podjedinica. Takve interakcije su važne za smanjenje podjedinica organiziranog kompleksa. Dionice podjedinica koje međusobno djeluju nazivaju se kontaktni majdani.
Klasičan primjer proteina koji ima četvrtinsku strukturu je hemoglobin. Molekula hemoglobina s molekularnom težinom od 68 000 sastoji se od četiri podjedinice dva različita tipa. α і β / α -Podjedinica se sastoji od 141 aminokiseline, a β - iz 146. Tercijarna struktura α - І β -podjedinica je slična svojoj molekularnoj težini (17 000 So). Podjedinica kože koja će se osvetiti protetskoj skupini - hem . Fragmenti hema prisutni su i u drugim proteinima (citokrom, mioglobin), koji nastaju dalje, ali želimo se ukratko osvrnuti na strukturu istih (slika 6). Grupirani hem je presavijeni koplanarni ciklički sustav koji se sastoji od središnjeg atoma, koji stvara koordinacijske veze s nekim viškom polimera, povezanih metanskim mjestima (= CH -). U hemoglobinu se počinje javljati razina oksidacije (2+).
Nekoliko podjedinica – dvije α i dva β - ujediniti u jedinstvenu strukturu na način da α -podjedinice samo kontakt β -podjedinice i kao cjelina (slika 7).

Mali 6. Građa hem hemoglobina

Mali 7. Shematski prikaz kvartarne strukture hemoglobina:
Fe - hem hemoglobin
Kao što se može vidjeti kod bebe 7, jedna molekula hemoglobina sposobna je prenijeti 4 molekule kiseline. I vezanje i otpuštanje kiselosti praćeni su konformacijskim promjenama u strukturi α - І β -podjedinica hemoglobina i njihovo međusobno otapanje u epimolekuli Ova činjenica potvrđuje da četvrtinska struktura proteina više nije apsolutno kruta.
Slične informacije.
Jedna od značajki proteina je njihova složena strukturna organizacija. Svi proteini imaju primarnu, sekundarnu i tercijarnu strukturu, a oni koji imaju dva ili više PPC imaju kvaternarnu strukturu (QS).
Primarna struktura proteina (PSB) – Ovo je redoslijed taloženja (slijed) viškova aminokiselina u PPC.
Međutim, zbog svoje zastupljenosti i sastava aminokiselina, proteini mogu biti različitih vrsta. Na primjer, iz dvije aminokiseline možete kombinirati 2 različita dipeptida:
S brojem aminokiselina većim od 20, broj mogućih kombinacija manji je od 210 18 . A ako znate da u PPC aminokiselina kože može postati osjetljiva više od jednom, onda je važno zaštititi broj mogućih opcija.
Značenje primarne strukture proteina (PPS).
Za pomoć se može izračunati PSB proteina feniltiohidantoin
metoda
. Ova se metoda temelji na međusobnoj reakciji fenilizotiocijanat
(FITC) s α-AA. Kao rezultat toga, stvara se kompleks dvaju odnosa FITC-AK
.
Na primjer, pogledajmo peptid ![]() Pomoću ove metode, PSB se koristi za određivanje slijeda viškova aminokiselina.
Pomoću ove metode, PSB se koristi za određivanje slijeda viškova aminokiselina.
FITC stupa u interakciju s terminalnom aminokiselinom (a). Kompleks je u osnivanju FTG-a, koji se dodaje u smjesu i ukazuje na snagu aminokiseline. A. Na primjer, tse - asn
![]() itd. Dosljedno jačati i identificirati sve ostale aminokiseline. To je naporan proces. PSB protein srednje veličine traje nekoliko mjeseci.
itd. Dosljedno jačati i identificirati sve ostale aminokiseline. To je naporan proces. PSB protein srednje veličine traje nekoliko mjeseci.
Dešifriranje PSB-a ima prioritet Sengeru(1953), što je otkriće PSB inzulina (dobitnik Nobelove nagrade). Molekula inzulina sastoji se od 2 PPC-a – A i B.

A-lanceta se sastoji od 21 aminokiseline, lanceta - od 30. PPC su međusobno povezani disulfidnim mjestima. Broj proteina čiji su PSB identificirani danas doseže 1500. Manje promjene u primarnoj strukturi mogu značajno promijeniti snagu proteina. U eritrocitima zdravih ljudi HbA je prisutan - kada je zamijenjen u -lanceru HbA, na 6. poziciji glu na vratilo krivi za tešku bolest anemija srpastih stanica, jer sva djeca rođena s ovom anomalijom umiru rano. S druge strane, postoje moguće opcije za promjenu PSB-a, koje ne navode fizikalno-kemijski i biološki autoriteti. Na primjer, HbC je na 6. poziciji b-koplja umjesto gluliza, HbC možda neće konkurirati svojom snagom nad HbA, a ljudi koji nose takav Hb u svojim eritrocitima su praktički zdravi.
Stabilnost PSB-a Osiguran je uglavnom unutarnjim kovalentnim peptidnim vezama i, drugim riječima, disulfidnim vezama.
Sekundarna struktura proteina (PSB).
PPV proteina pokazuje veliku fleksibilnost i razvija izrazitu prostranu strukturu ili konformacija. U proteinima postoje 2 razine takve konformacije - to je VSB i tercijarna struktura (TBB).
VSB – Ova konfiguracija PPC-a, način na koji je položen ili uvijen u bilo koju konformaciju, u skladu je s programom sadržanim u P SB.
Postoje tri glavne vrste VSB:
1) -spirala;
2) b-struktura(kugla zaliha dijelova ili presavijeni list s dijelovima);
3) lopta bez pragova.
-spirala .
Ovaj model predložio je V. Pauling. Ovo je najbolje za globularne proteine. Za svaki sustav najstabilnije stanje je ono koje pokazuje minimum slobodne energije. Za peptide se ova situacija događa kada su CO- i NH-skupine međusobno povezane slabom vodenom vezom. U a -spirale NH skupina 1. aminokiselinskog ostatka u interakciji je s CO skupinom 4. aminokiseline. Kao rezultat toga, okosnica peptida stvara spiralu, na kojoj višak od 3,6 AA pada na kožni okret.
1 spiralna spirala (1 zavoj) = 3,6 AC = 0,54 nm, rez prema gore – 26°
Uvijanje PPC-a promatra se iza strelice godine, tako da se spirala pomiče udesno. Kroz kožu 5 zavoja (18 AC; 2,7 nm) PPC konfiguracija se ponavlja.
Stabilizirajući VSB prvo prije vodenih veza, au drugom - peptidne i disulfidne. Vodene veze su 10-100 puta slabije od standardnih kemijskih veza; međutim, njihova velika količina smrada osigurat će oštrinu i kompaktnost VSB-a. R-lancele su a-spiralne, skošene do točke prstena i raširene duž različitih strana i osi.
b -struktura .
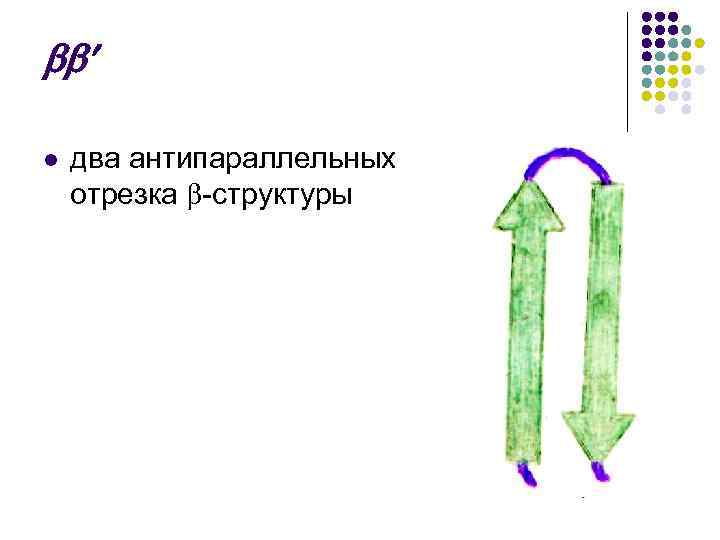
To su presavijeni dijelovi PPC parcele, koji podsjećaju na oblik lista, presavijeni poput harmonike. Lopte PPC-a mogu biti paralelne, budući da napad počinje na N- ili C-kraju.
Budući da su uska koplja lopte usmjerena s proksimalnim krajevima N–C i C–N, tada se smrdi nazivaju antiparalelan.

 paralelno
paralelno
![]() antiparalelan
antiparalelan
Stvaranje vodenih veza događa se, kao u a-heliksu, između CO i NH skupina.

 l l Proučavanje strukturne organizacije proteina jedan je od glavnih problema suvremene biokemije.Ima važno znanstveno i praktično značenje za razumijevanje velike ekspanzije funkcija proteina. Postoje li deseci ili stotine aminokiselina u proteinskoj molekuli?
l l Proučavanje strukturne organizacije proteina jedan je od glavnih problema suvremene biokemije.Ima važno znanstveno i praktično značenje za razumijevanje velike ekspanzije funkcija proteina. Postoje li deseci ili stotine aminokiselina u proteinskoj molekuli?
 Emil Fisher Proteini su presavijeni polipeptidi u kojima su aminokiseline povezane peptidnim vezama (R-CO-NH-R) koje nastaju tijekom interakcije karboksilnih i amino skupina aminokiselina.
Emil Fisher Proteini su presavijeni polipeptidi u kojima su aminokiseline povezane peptidnim vezama (R-CO-NH-R) koje nastaju tijekom interakcije karboksilnih i amino skupina aminokiselina.
 Eksperimentalni dokaz teorije l l l Prirodni proteini imaju mali broj titriranih skupina – COOH i –NH 2. Tijekom procesa hidrolize proteina pod djelovanjem proteolita stvara se stehiometrijski broj titriranih skupina – COOH i –NH 2. U prisutnosti odreenih enzima proteini se razgrađuju na strogo određene fragmente (polipeptide).RTG analiza potvrđuje strukturu polipeptida Protein Glavna potvrda polipeptidne teorije proteina je mogućnost sintetiziranja polipeptida i proteina iz već poznatih. protein (inzulin, lizozim, ribonukleus) kemijskim metodama.
Eksperimentalni dokaz teorije l l l Prirodni proteini imaju mali broj titriranih skupina – COOH i –NH 2. Tijekom procesa hidrolize proteina pod djelovanjem proteolita stvara se stehiometrijski broj titriranih skupina – COOH i –NH 2. U prisutnosti odreenih enzima proteini se razgrađuju na strogo određene fragmente (polipeptide).RTG analiza potvrđuje strukturu polipeptida Protein Glavna potvrda polipeptidne teorije proteina je mogućnost sintetiziranja polipeptida i proteina iz već poznatih. protein (inzulin, lizozim, ribonukleus) kemijskim metodama.

 Strukturne značajke peptidnog koplja l l Peptidna veza je približno 10% kraća od –C–N– veze i ima karakter „često uronjene” veze –C=N– prema L. Paulingu i R. Coreyu, koja je bila razvio se 1948–1955 pp. objasniti posebnu prirodu C–N veze "rezonancijom" između dva oblika
Strukturne značajke peptidnog koplja l l Peptidna veza je približno 10% kraća od –C–N– veze i ima karakter „često uronjene” veze –C=N– prema L. Paulingu i R. Coreyu, koja je bila razvio se 1948–1955 pp. objasniti posebnu prirodu C–N veze "rezonancijom" između dva oblika
 Strukturne značajke peptidnog koplja l l l Drugim riječima, u proteinima i peptidima C–N veze su često višestruke kroz interakciju usamljenog para elektrona atoma dušika s elektroničkim sustavom karbonilne skupine, što dovodi do teškog formacija tannya navkolo zvjazku C–N Zazvichay peptid zv' Trans konfiguracija je izraženija od cis konfiguracije za 2,6 kcal/mol (10,878 kJ/mol), budući da blizina ugljikovih atoma u cis konfiguraciji otežava
Strukturne značajke peptidnog koplja l l l Drugim riječima, u proteinima i peptidima C–N veze su često višestruke kroz interakciju usamljenog para elektrona atoma dušika s elektroničkim sustavom karbonilne skupine, što dovodi do teškog formacija tannya navkolo zvjazku C–N Zazvichay peptid zv' Trans konfiguracija je izraženija od cis konfiguracije za 2,6 kcal/mol (10,878 kJ/mol), budući da blizina ugljikovih atoma u cis konfiguraciji otežava
 Strukturne značajke peptidnog koplja l l l Moguće su rotacije oko dvije jednostavne veze (N–C i C–C 1), koje prianjaju na asimetrični atom = 180 - trans-forma)
Strukturne značajke peptidnog koplja l l l Moguće su rotacije oko dvije jednostavne veze (N–C i C–C 1), koje prianjaju na asimetrični atom = 180 - trans-forma)
 Strukturne značajke peptidnog lanceta l l Zbog međudjelovanja između branitelja u peptidnom lancetu, oni ne mogu poprimiti nikakvo značenje - dopuštena su im samo određena diskretna područja koja ukazuju na energetsku snagu Neke konformacije peptida lanjug Kuti imaju međusobne odnose, promjena jednog od njih je teža promjena drugog
Strukturne značajke peptidnog lanceta l l Zbog međudjelovanja između branitelja u peptidnom lancetu, oni ne mogu poprimiti nikakvo značenje - dopuštena su im samo određena diskretna područja koja ukazuju na energetsku snagu Neke konformacije peptida lanjug Kuti imaju međusobne odnose, promjena jednog od njih je teža promjena drugog

 l K. W. Linderström-Lang razlikovao je 4 razine organizacije proteinskih molekula - primarnu, sekundarnu, tercijarnu i kvartarnu strukturu. Iako su ove kategorije zastarjele, zasad će se nastaviti koristiti.
l K. W. Linderström-Lang razlikovao je 4 razine organizacije proteinskih molekula - primarnu, sekundarnu, tercijarnu i kvartarnu strukturu. Iako su ove kategorije zastarjele, zasad će se nastaviti koristiti.

 Primarna struktura proteina l l l sekvenca aminokiselinskih ostataka u polipeptidnoj lanceti kodirana je strukturnim genom ovog proteina i sadrži sve što je potrebno za samoorganizaciju njegove prostrane strukture. Svi proteini se natječu za vlastitu S ovom primarnom strukturom, potencijalni broj takvih struktura nije ograničen na broj različitih vrsta proteina u svim vrstama živih organizama oko 1010-1012
Primarna struktura proteina l l l sekvenca aminokiselinskih ostataka u polipeptidnoj lanceti kodirana je strukturnim genom ovog proteina i sadrži sve što je potrebno za samoorganizaciju njegove prostrane strukture. Svi proteini se natječu za vlastitu S ovom primarnom strukturom, potencijalni broj takvih struktura nije ograničen na broj različitih vrsta proteina u svim vrstama živih organizama oko 1010-1012
 l l l Nemoguće je razumjeti biološku funkciju i molekularni mehanizam fiziološke aktivnosti proteina bez detaljnog poznavanja njegovog postojanja. Istraživanje primarne strukture "mutagenih" proteina omogućuje razumijevanje prirode recesijskih bolesti na molekularnoj razini.
l l l Nemoguće je razumjeti biološku funkciju i molekularni mehanizam fiziološke aktivnosti proteina bez detaljnog poznavanja njegovog postojanja. Istraživanje primarne strukture "mutagenih" proteina omogućuje razumijevanje prirode recesijskih bolesti na molekularnoj razini.
 Metode određivanja primarne strukture l l Priprema proteina prije analize primarne strukture ima za cilj minimizirati infuziju drugih, viših razina njegove organizacije. Drugim riječima, predmet analize može biti poremećeno ovo proteinsko koplje bez ikakvih poprečne kovalentne veze (na primjer, disulfid) tako da su sve peptidne veze dostupne i kemijskim reagensima i enzimima
Metode određivanja primarne strukture l l Priprema proteina prije analize primarne strukture ima za cilj minimizirati infuziju drugih, viših razina njegove organizacije. Drugim riječima, predmet analize može biti poremećeno ovo proteinsko koplje bez ikakvih poprečne kovalentne veze (na primjer, disulfid) tako da su sve peptidne veze dostupne i kemijskim reagensima i enzimima
 l l Protein je prvo odgovoran za svu duboku denaturaciju i gubitak kvaternarne, tercijarne i, ako je moguće, sekundarne strukture.
l l Protein je prvo odgovoran za svu duboku denaturaciju i gubitak kvaternarne, tercijarne i, ako je moguće, sekundarne strukture.
 l Kako biste spriječili stvaranje disulfidnih veza, ponovno blokirajte sulfhidrilne skupine viškom jodoktične kiseline: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI
l Kako biste spriječili stvaranje disulfidnih veza, ponovno blokirajte sulfhidrilne skupine viškom jodoktične kiseline: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI
 Zatim se provodi selektivna hidroliza denaturiranog proteina kako bi se uklonio peptidni sustav koji se preklapa. Hidroliza se provodi enzimima: l l l tripsin (veze stvaraju karboksilne skupine aromatskih aminokiselina), kimotripsin (veze stvaraju -COOH skupine aromatskih aminokiselina) i dr.; kemijski reagensi: bromcijan Br - C N (vezivna sredstva, složena sa skupinama - COOH met) i dr.
Zatim se provodi selektivna hidroliza denaturiranog proteina kako bi se uklonio peptidni sustav koji se preklapa. Hidroliza se provodi enzimima: l l l tripsin (veze stvaraju karboksilne skupine aromatskih aminokiselina), kimotripsin (veze stvaraju -COOH skupine aromatskih aminokiselina) i dr.; kemijski reagensi: bromcijan Br - C N (vezivna sredstva, složena sa skupinama - COOH met) i dr.
 l l l Prva faza - frakcioniranje izoliranih peptida - provodi se Nashovim kromatografskim metodama, a preostala faza - određivanje primarne strukture l počinje od značaja molekulske mase, sastava aminokiselina, N- i terminalnih aminokiselina u višku. drugo sredstvo i odaberite drugu, različitu vrstu iz prvog skupa peptidnih fragmenata, koji se dijele i analiziraju kako bi se odredio redoslijed distribucije aminokiselina u proteinu
l l l Prva faza - frakcioniranje izoliranih peptida - provodi se Nashovim kromatografskim metodama, a preostala faza - određivanje primarne strukture l počinje od značaja molekulske mase, sastava aminokiselina, N- i terminalnih aminokiselina u višku. drugo sredstvo i odaberite drugu, različitu vrstu iz prvog skupa peptidnih fragmenata, koji se dijele i analiziraju kako bi se odredio redoslijed distribucije aminokiselina u proteinu

 F. Sangerova metoda l U 1945 r. Engleski biokemičar F. Sanger razvio je jednu od prvih metoda za identifikaciju N-terminalnih aminokiselina
F. Sangerova metoda l U 1945 r. Engleski biokemičar F. Sanger razvio je jednu od prvih metoda za identifikaciju N-terminalnih aminokiselina
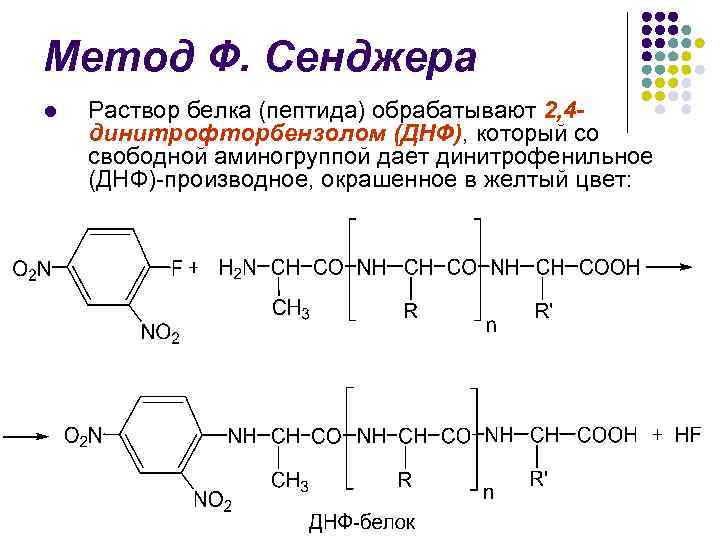 Metoda F. Sangera l Protein (peptid) se tretira s 2,4 dinitrofluorobenzenom (DNF), koji iz slobodne amino skupine daje dinitrofenil (DNF) koji sadrži žutu boju:
Metoda F. Sangera l Protein (peptid) se tretira s 2,4 dinitrofluorobenzenom (DNF), koji iz slobodne amino skupine daje dinitrofenil (DNF) koji sadrži žutu boju:
 Metoda F. Sangera l l Napadna kiselinska hidroliza (5,7 N HCl) dovodi do razaranja peptidnih veza i stvaranja DNP-like N-terminalne aminokiseline.
Metoda F. Sangera l l Napadna kiselinska hidroliza (5,7 N HCl) dovodi do razaranja peptidnih veza i stvaranja DNP-like N-terminalne aminokiseline.
 Dansilny metoda l Rozrobleny 1963. godine. engleski biokemičari W. Gray i B. Hartley
Dansilny metoda l Rozrobleny 1963. godine. engleski biokemičari W. Gray i B. Hartley
 Dansil metoda l Prvi stupanj - reakcija dansil klorida (1-dimetilaminonaftalen-5-sulfoklorida) s neprotoniranim peptidom (ili proteinom) amino skupine s modificiranim dansil peptidom (DNS peptid)
Dansil metoda l Prvi stupanj - reakcija dansil klorida (1-dimetilaminonaftalen-5-sulfoklorida) s neprotoniranim peptidom (ili proteinom) amino skupine s modificiranim dansil peptidom (DNS peptid)

 Najjača metoda uklanjanja DNS proteina je hidroliza u prisutnosti 5, 7 n. HCl, na 105 Tijekom razdoblja od 12-16 godina, nakon čega je DNS-aminokiselina odvojena, identificirana fluorescencijom u UV-izmjenama
Najjača metoda uklanjanja DNS proteina je hidroliza u prisutnosti 5, 7 n. HCl, na 105 Tijekom razdoblja od 12-16 godina, nakon čega je DNS-aminokiselina odvojena, identificirana fluorescencijom u UV-izmjenama


 Metoda S. Akabori l l Kada se peptid (ili protein) zagrijava s bezvodnim hidrazinom na 100-120 C, peptidne veze se hidroliziraju s aminokiselinama hidrazida, C-terminalna aminokiselina gubi izgled Ostale aminokiseline mogu vidjeti iz smjese i identificirati:
Metoda S. Akabori l l Kada se peptid (ili protein) zagrijava s bezvodnim hidrazinom na 100-120 C, peptidne veze se hidroliziraju s aminokiselinama hidrazida, C-terminalna aminokiselina gubi izgled Ostale aminokiseline mogu vidjeti iz smjese i identificirati:

 Metoda P. Edmana l Metodu razgradnje polipeptidnog lanceta uz pomoć fenilizotiocionata (FITC) razradio je švedski kemičar P. V. Edman 1950-56. l Edmanova metoda omogućuje dosljedno uklanjanje N-terminalnih aminokiselinskih ostataka iz oblika feniltiohidantoina (PTH) l Ciklus razgradnje kože uključuje 3 faze
Metoda P. Edmana l Metodu razgradnje polipeptidnog lanceta uz pomoć fenilizotiocionata (FITC) razradio je švedski kemičar P. V. Edman 1950-56. l Edmanova metoda omogućuje dosljedno uklanjanje N-terminalnih aminokiselinskih ostataka iz oblika feniltiohidantoina (PTH) l Ciklus razgradnje kože uključuje 3 faze
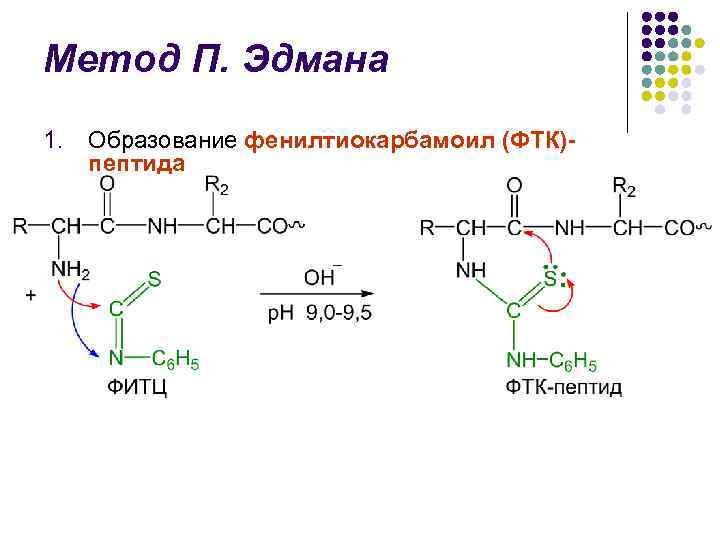



 P. Edmanova metoda l l l Identifikacija PTG separacija je inicijalno u Edmanovoj metodi. Tijekom posljednja tri sata provedena je kromatografija na papiru, zatim mikrofina sferna kromatografija na silika gelu i poliamidu, izvorna kromatografija prirodnog plina Velika postignuća u području strukturne istraživanje. Edman i J. Begg sekvencer (od engl. sequence - slijed) - uređaj koji uz visoku učinkovitost automatski odvaja N-terminalne aminokiselinske ostatke Edmanovom metodom.
P. Edmanova metoda l l l Identifikacija PTG separacija je inicijalno u Edmanovoj metodi. Tijekom posljednja tri sata provedena je kromatografija na papiru, zatim mikrofina sferna kromatografija na silika gelu i poliamidu, izvorna kromatografija prirodnog plina Velika postignuća u području strukturne istraživanje. Edman i J. Begg sekvencer (od engl. sequence - slijed) - uređaj koji uz visoku učinkovitost automatski odvaja N-terminalne aminokiselinske ostatke Edmanovom metodom.
 l Za određivanje strukture peptida i proteina mogu se sažeti: l l enzimi koji kataliziraju eliminaciju N- i C-terminalnih aminokiselinskih ostataka - amino karboksipeptidaze, fizikalno-kemijske metode, spektroskopija, masena spektrometrija yu
l Za određivanje strukture peptida i proteina mogu se sažeti: l l enzimi koji kataliziraju eliminaciju N- i C-terminalnih aminokiselinskih ostataka - amino karboksipeptidaze, fizikalno-kemijske metode, spektroskopija, masena spektrometrija yu
 Analiza podataka o primarnoj strukturi omogućuje razvoj daljnjih osnovnih principa Stabilnost primarne strukture osiguravaju uglavnom peptidne veze; Moguće sudjelovanje i mali broj disulfidnih veza U polipeptidnom lancu mogu se otkriti različite kombinacije aminokiselina U polipeptidima se otkrivaju svi mogući dipeptidi.
Analiza podataka o primarnoj strukturi omogućuje razvoj daljnjih osnovnih principa Stabilnost primarne strukture osiguravaju uglavnom peptidne veze; Moguće sudjelovanje i mali broj disulfidnih veza U polipeptidnom lancu mogu se otkriti različite kombinacije aminokiselina U polipeptidima se otkrivaju svi mogući dipeptidi.
 Analiza podataka o primarnoj strukturi omogućuje nam da razvijemo sljedeće osnovne principe: l l Kožu pojedinog homogenog proteina karakterizira jedinstvena primarna struktura; Često zamjena aminokiselina dovodi ne samo do strukturnih promjena, već i do promjena u fizičkim i kemijskim svojstvima i biološkim funkcijama. Ovaj princip strukturne sličnosti je najtipičniji za niske proteolitičke enzime - tripsin, kimotripsin itd.
Analiza podataka o primarnoj strukturi omogućuje nam da razvijemo sljedeće osnovne principe: l l Kožu pojedinog homogenog proteina karakterizira jedinstvena primarna struktura; Često zamjena aminokiselina dovodi ne samo do strukturnih promjena, već i do promjena u fizičkim i kemijskim svojstvima i biološkim funkcijama. Ovaj princip strukturne sličnosti je najtipičniji za niske proteolitičke enzime - tripsin, kimotripsin itd.

 U nepolarnoj fuziji, energija vodenog ligamenta –CO NN– postaje blizu 16,7 kJ/mol, a pomak u polaritetu srednje struje smanjuje tu energiju
U nepolarnoj fuziji, energija vodenog ligamenta –CO NN– postaje blizu 16,7 kJ/mol, a pomak u polaritetu srednje struje smanjuje tu energiju
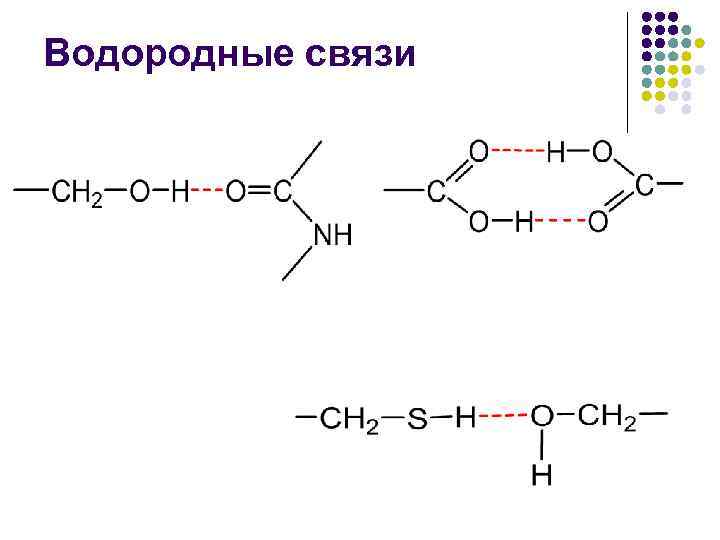
 Hidrofobne interakcije imaju entropijsku prirodu zbog činjenice da nepolarni branitelji ulaze u vodu i pokušavaju uspostaviti kontakt s njom.Na primjer, voda ne obnavlja svoju strukturnu strukturu i kao da je Primus grupa. klaster koji sadrži minimalnu energiju. višak
Hidrofobne interakcije imaju entropijsku prirodu zbog činjenice da nepolarni branitelji ulaze u vodu i pokušavaju uspostaviti kontakt s njom.Na primjer, voda ne obnavlja svoju strukturnu strukturu i kao da je Primus grupa. klaster koji sadrži minimalnu energiju. višak

 Van der Waalsove interakcije l l Sastoje se od disperzijskih sila, gravitacije atoma i sila međusobne prilagodbe njihovih elektroničkih ljuski. Doprinos energije kontaktu s kožom je mali (
Van der Waalsove interakcije l l Sastoje se od disperzijskih sila, gravitacije atoma i sila međusobne prilagodbe njihovih elektroničkih ljuski. Doprinos energije kontaktu s kožom je mali (
 Ionske ili elektrostatske interakcije l Interakcije nabijenih skupina l l Interakcije ionogenih skupina koje stvaraju solne veze Energija solnih veza može doseći hidrofobnu površinu od 41,9 k.J/mol, ali njihova je količina dosta mala. slične ion-dipolne i dipol-dipolne interakcije
Ionske ili elektrostatske interakcije l Interakcije nabijenih skupina l l Interakcije ionogenih skupina koje stvaraju solne veze Energija solnih veza može doseći hidrofobnu površinu od 41,9 k.J/mol, ali njihova je količina dosta mala. slične ion-dipolne i dipol-dipolne interakcije
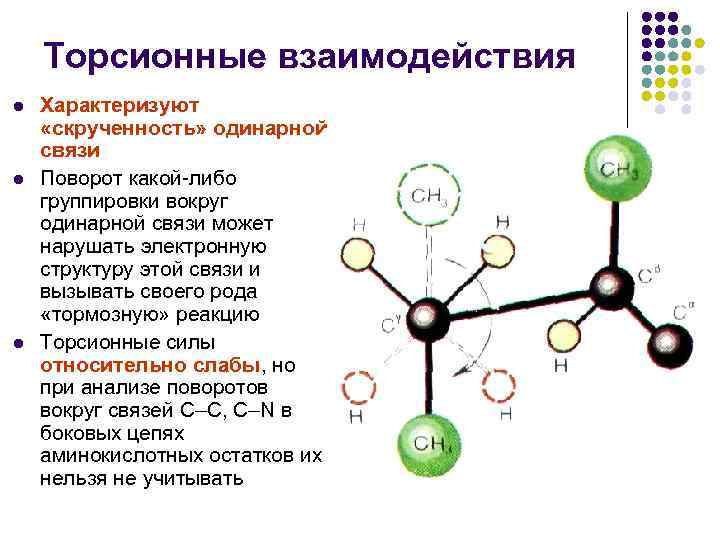 Torzijske interakcije l l l Karakteriziraju "uvijanje" jednostruke veze. Rotacija bilo koje grupe oko jednostruke veze može uništiti elektroničku strukturu njezine veze i pokrenuti neku vrstu "halm" reakcije. Torzijska reakcija i sile očito su slabe, ali kada analizirajući zavoje oko ligamenata íS, íN otpadnih lanceta ne može se izbjeći višak aminokiselina
Torzijske interakcije l l l Karakteriziraju "uvijanje" jednostruke veze. Rotacija bilo koje grupe oko jednostruke veze može uništiti elektroničku strukturu njezine veze i pokrenuti neku vrstu "halm" reakcije. Torzijska reakcija i sile očito su slabe, ali kada analizirajući zavoje oko ligamenata íS, íN otpadnih lanceta ne može se izbjeći višak aminokiselina

 Sekundarna struktura proteina... l l ekstenzivno širenje zaobljenih dijelova polipeptidne lancete bez vrste i konformacije radikala aminokiselina.Nastaje interakcijom vodenih veza između peptidnih skupina kako jedne tako i različitih lanceta. Lantsyugiv
Sekundarna struktura proteina... l l ekstenzivno širenje zaobljenih dijelova polipeptidne lancete bez vrste i konformacije radikala aminokiselina.Nastaje interakcijom vodenih veza između peptidnih skupina kako jedne tako i različitih lanceta. Lantsyugiv
 Sekundarna struktura proteina l l l Svaki dio proteinske molekule ima sekundarnu strukturu. Neki smatraju sekundarnom strukturom periodičnije elemente: - spiralu i - strukturu. Međutim, u proteinima dijelovi, koji su strukturirani, postaju precizniji u nježan način, iako svojom prostranom strukturom ne osvećuje nikoga s vremena na vrijeme ponovljeni, pravilni motiv. Prije njih, u potpunosti razumijemo koncept sekundarne strukture. Postoje 2 vrste sekundarnih struktura: pravilne i nepravilne. Koncept sekundarne strukture ne odnosi se na sve proteinske molekule kao cjelinu, već na nekoliko većih dijelova poda. ipeptidna lanceta
Sekundarna struktura proteina l l l Svaki dio proteinske molekule ima sekundarnu strukturu. Neki smatraju sekundarnom strukturom periodičnije elemente: - spiralu i - strukturu. Međutim, u proteinima dijelovi, koji su strukturirani, postaju precizniji u nježan način, iako svojom prostranom strukturom ne osvećuje nikoga s vremena na vrijeme ponovljeni, pravilni motiv. Prije njih, u potpunosti razumijemo koncept sekundarne strukture. Postoje 2 vrste sekundarnih struktura: pravilne i nepravilne. Koncept sekundarne strukture ne odnosi se na sve proteinske molekule kao cjelinu, već na nekoliko većih dijelova poda. ipeptidna lanceta
 Sekundarna struktura proteina l Interakcije koje igraju najvažniju ulogu u formiranoj sekundarnoj strukturi označene su: l l l karakteristikama peptidne veze i prostornim interakcijama (- i -rezovi) Glavnu ulogu imaju vodene veze između peptidne skupine, koje se periodički ponavljaju u Lancusu
Sekundarna struktura proteina l Interakcije koje igraju najvažniju ulogu u formiranoj sekundarnoj strukturi označene su: l l l karakteristikama peptidne veze i prostornim interakcijama (- i -rezovi) Glavnu ulogu imaju vodene veze između peptidne skupine, koje se periodički ponavljaju u Lancusu
 Stabilizacija sekundarne strukture za stvaranje veziva vode l l Ubrizgavanje viška proteina s vodom Voda se može učinkovito natjecati za stvaranje veziva vode: Infuzija vode se smanjuje kada se oblikuje kompaktna prostrana proteinska struktura, rast i umjesto peptidnih veza, povećana kompatibilnost njihovih međudjelovanja Dakle, stabilnost sekundarne strukture leži u kompaktnoj tercijarnoj strukturi
Stabilizacija sekundarne strukture za stvaranje veziva vode l l Ubrizgavanje viška proteina s vodom Voda se može učinkovito natjecati za stvaranje veziva vode: Infuzija vode se smanjuje kada se oblikuje kompaktna prostrana proteinska struktura, rast i umjesto peptidnih veza, povećana kompatibilnost njihovih međudjelovanja Dakle, stabilnost sekundarne strukture leži u kompaktnoj tercijarnoj strukturi

 -Spirala l U 50-im stijenama XX. stoljeća. L. Pauling i R. Corey, na temelju podataka o strukturi kristala aminokiselina i jednostavnih peptida, promatrali su moguće periodične konformacije polipeptida lanjuga i došli do zaključka da su najhomogeniju strukturu po njima nazvali -spi rally.
-Spirala l U 50-im stijenama XX. stoljeća. L. Pauling i R. Corey, na temelju podataka o strukturi kristala aminokiselina i jednostavnih peptida, promatrali su moguće periodične konformacije polipeptida lanjuga i došli do zaključka da su najhomogeniju strukturu po njima nazvali -spi rally.
 -Spirala Odabir postavljanja temelji se na sljedećim kriterijima: 1. Stvaranje čvrsto zbijene kompaktne strukture bez praznih i pretjerane kritike atoma 2. Maksimalna zasićenost strukture vodenim vezama iz ove kemikalije, tako da je njihova geometrija bliska linearnoj 3 .. Dotrimannya između atomskih uspona i kutova i oprostite peptidima l
-Spirala Odabir postavljanja temelji se na sljedećim kriterijima: 1. Stvaranje čvrsto zbijene kompaktne strukture bez praznih i pretjerane kritike atoma 2. Maksimalna zasićenost strukture vodenim vezama iz ove kemikalije, tako da je njihova geometrija bliska linearnoj 3 .. Dotrimannya između atomskih uspona i kutova i oprostite peptidima l
 -Spirala l U glavama ovih ljudi može postojati i desna i lijeva spirala, ali za desnu, spirala se čini energetski istaknutija od lijeve, budući da je peptidno koplje stvoreno s L-aminokiselinama
-Spirala l U glavama ovih ljudi može postojati i desna i lijeva spirala, ali za desnu, spirala se čini energetski istaknutija od lijeve, budući da je peptidno koplje stvoreno s L-aminokiselinama
 -Spiralna l l Spiralna struktura se razvija ako se sve strane polipeptidnog koplja okreću oko jednostavnih ligamenata (i) imaju istu vrijednost i predznak (blizu 60), što dovodi do postupnog uvijanja koplja u kojem u slučaju da je radikala previše, kiseline se pojavljuju na periferiji spirale otopljenog cilindra i mogu, ovisno o svojoj prirodi, osigurati hidrofobnu ili hidrofilnu prirodu cilindrične površine
-Spiralna l l Spiralna struktura se razvija ako se sve strane polipeptidnog koplja okreću oko jednostavnih ligamenata (i) imaju istu vrijednost i predznak (blizu 60), što dovodi do postupnog uvijanja koplja u kojem u slučaju da je radikala previše, kiseline se pojavljuju na periferiji spirale otopljenog cilindra i mogu, ovisno o svojoj prirodi, osigurati hidrofobnu ili hidrofilnu prirodu cilindrične površine
 -Geometrijski parametri spirale: l l l l radijus r = 2,3 Å (0,23 nm) visina spirale (pomak) po 1 višku d = 1,5 Å rub spirale (period identiteta) P = 5,4 Å 1 zavoj spirale, oblik. 3, 6 višak aminokiselina, sve – Veze C=O su ravno naprijed, do C-kraja, a skupine –N–H su ravno natrag.U spirali je kožna skupina –NH povezana vodenom vezom skupini –CO četvrte aminokiseline previše. znyogo (5 1 poziv)
-Geometrijski parametri spirale: l l l l radijus r = 2,3 Å (0,23 nm) visina spirale (pomak) po 1 višku d = 1,5 Å rub spirale (period identiteta) P = 5,4 Å 1 zavoj spirale, oblik. 3, 6 višak aminokiselina, sve – Veze C=O su ravno naprijed, do C-kraja, a skupine –N–H su ravno natrag.U spirali je kožna skupina –NH povezana vodenom vezom skupini –CO četvrte aminokiseline previše. znyogo (5 1 poziv)
 -Spirala l l Broj -spiralnih sekcija kod globularnih proteina je izuzetno mali (5 - 15 viška aminokiselina, 3 - 4 zavoja spirale), kod fibrilarnih proteina - bogato proširen.Molim vas nazovite na mjestima gdje je uključen višak tako da se sustavi mogu prekinuti. Tada se cijela spirala skuplja za 20-30
-Spirala l l Broj -spiralnih sekcija kod globularnih proteina je izuzetno mali (5 - 15 viška aminokiselina, 3 - 4 zavoja spirale), kod fibrilarnih proteina - bogato proširen.Molim vas nazovite na mjestima gdje je uključen višak tako da se sustavi mogu prekinuti. Tada se cijela spirala skuplja za 20-30

 -Struktura l l Predložio W. T. Astbury 1941. godine. na temelju rendgenskih strukturnih studija - keratin Nakon 10 godina, L. Pauling i R. Corey ustanovili su da je struktura, ili "preklopna ploča", posljedica stabilizacije međulancinarnih vodenih ligamenata koji povezuju pleteni, cik-cak peptidni ligamenti
-Struktura l l Predložio W. T. Astbury 1941. godine. na temelju rendgenskih strukturnih studija - keratin Nakon 10 godina, L. Pauling i R. Corey ustanovili su da je struktura, ili "preklopna ploča", posljedica stabilizacije međulancinarnih vodenih ligamenata koji povezuju pleteni, cik-cak peptidni ligamenti




 -Struktura l l Broj viška aminokiselina u dijelu lanceleta peptida, koji stvara -strukturu, kreće se od 3 do 8. Proširena struktura, tzv. lopta, ili presavijeni list, najčešće se sastoji od 2- 6 lanceta, ponekad i do 10
-Struktura l l Broj viška aminokiselina u dijelu lanceleta peptida, koji stvara -strukturu, kreće se od 3 do 8. Proširena struktura, tzv. lopta, ili presavijeni list, najčešće se sastoji od 2- 6 lanceta, ponekad i do 10
 -Struktura l l l Kisele skupine viška aminokiselina, kada se stvori struktura, otkrivaju se različitim stranama svoje površine. Sama površina ima naboran oblik, a nabori su definirani atomima ugljika. Organske skupine, koje dolaze iz njih, nastaju češalj. To vam omogućuje oblikovanje i produživanje dugih površina ispunjenih istom vrstom (na primjer, hidrofobnih) bioloških radikala hidrofobne površine presavijene lopte, u interakciji jedna s drugom ili s hidrofobnim grebenima spirala , sudjeluju u unutarnjim molekularnim hidrofobnim jezgrama, što je lako stabilizirati strukturu proteina
-Struktura l l l Kisele skupine viška aminokiselina, kada se stvori struktura, otkrivaju se različitim stranama svoje površine. Sama površina ima naboran oblik, a nabori su definirani atomima ugljika. Organske skupine, koje dolaze iz njih, nastaju češalj. To vam omogućuje oblikovanje i produživanje dugih površina ispunjenih istom vrstom (na primjer, hidrofobnih) bioloških radikala hidrofobne površine presavijene lopte, u interakciji jedna s drugom ili s hidrofobnim grebenima spirala , sudjeluju u unutarnjim molekularnim hidrofobnim jezgrama, što je lako stabilizirati strukturu proteina

 -Vigin l l I spirala i struktura predstavljeni su u globularnim proteinima u kratkim dijelovima, što znači da dio sekundarne strukture proteina pada na različite vrste petlji, što vam omogućuje promjenu smjera lanjuga peptida. Ekonomičan strukturni element koji omogućuje rotaciju polipeptida za 180 stupnjeva. 3 peptidne skupine, - -vigin Stabiliziran jednom vodenom vezom Gotovo uvijek se pojavljuje na površini proteinske globule, koja često igra važnu ulogu u njezinim interakcijama s drugim molekulama
-Vigin l l I spirala i struktura predstavljeni su u globularnim proteinima u kratkim dijelovima, što znači da dio sekundarne strukture proteina pada na različite vrste petlji, što vam omogućuje promjenu smjera lanjuga peptida. Ekonomičan strukturni element koji omogućuje rotaciju polipeptida za 180 stupnjeva. 3 peptidne skupine, - -vigin Stabiliziran jednom vodenom vezom Gotovo uvijek se pojavljuje na površini proteinske globule, koja često igra važnu ulogu u njezinim interakcijama s drugim molekulama


 l Rezultati rendgenske strukturne analize otkrili su da je način na koji je proteinska molekula savijena određen njezinim aminokiselinskim slijedom - gly, pro, asn Yakshcho sa 6 grupiranih viška aminokiselina 4 za kombiniranje stvaranja spirale, zatim ovu skupinu središte je spiralizacije. Ako se 3 dodatka od 5 grupiranih kombiniraju kako bi stvorili strukturu, tada je to grupiranje sjeme za
l Rezultati rendgenske strukturne analize otkrili su da je način na koji je proteinska molekula savijena određen njezinim aminokiselinskim slijedom - gly, pro, asn Yakshcho sa 6 grupiranih viška aminokiselina 4 za kombiniranje stvaranja spirale, zatim ovu skupinu središte je spiralizacije. Ako se 3 dodatka od 5 grupiranih kombiniraju kako bi stvorili strukturu, tada je to grupiranje sjeme za

 Sekundarna (suprasekundarna) struktura proteina l l Ova razina organizacije proteinskih molekula povezana je s prisutnošću skupova sekundarnih struktura koje međusobno djeluju.
Sekundarna (suprasekundarna) struktura proteina l l Ova razina organizacije proteinskih molekula povezana je s prisutnošću skupova sekundarnih struktura koje međusobno djeluju.

 Superspiralizirana -spirala l l Nastaje uvijanjem preslikača -spirala na protofibrilu (-keratin), koji se spaja u mikrofibril -Spirale su pritisnute jedna uz drugu Van der Waalsovim silama
Superspiralizirana -spirala l l Nastaje uvijanjem preslikača -spirala na protofibrilu (-keratin), koji se spaja u mikrofibril -Spirale su pritisnute jedna uz drugu Van der Waalsovim silama

 x l remen, koji se sastoji od dvije paralelne kuglice sa zglobovima između njih, izgleda: l neuredna kuglica - sa
x l remen, koji se sastoji od dvije paralelne kuglice sa zglobovima između njih, izgleda: l neuredna kuglica - sa
 x l traka, koja se sastoji od dvije paralelne kuglice sa zglobovima između njih, izgleda: l l - spirale - položena lanceta po Rossmanu Dva uzastopno spojena dijela stvaraju strukturu "grčkog ornamenta" -
x l traka, koja se sastoji od dvije paralelne kuglice sa zglobovima između njih, izgleda: l l - spirale - položena lanceta po Rossmanu Dva uzastopno spojena dijela stvaraju strukturu "grčkog ornamenta" -
 x l remen, koji se sastoji od dvije paralelne kuglice sa spojevima između njih, izgleda: l - struktura - - - cik-cak, “ključ od oraha”
x l remen, koji se sastoji od dvije paralelne kuglice sa spojevima između njih, izgleda: l - struktura - - - cik-cak, “ključ od oraha”

 domene i jedan po jedan kratki odsječak polipeptidnog koplja, koji se nazivaju zglobni odsječci. Funkcionalne domene mogu biti formirane od jedne ili više strukturnih domena. U nizu enzima aktivno središte je odvojeno između domena
domene i jedan po jedan kratki odsječak polipeptidnog koplja, koji se nazivaju zglobni odsječci. Funkcionalne domene mogu biti formirane od jedne ili više strukturnih domena. U nizu enzima aktivno središte je odvojeno između domena



 3. + -Proteini su dijelovi, uglavnom izgrađeni od spirala, i dijelovi, sastavljeni od anti-paralelnih kuglica Inzulin
3. + -Proteini su dijelovi, uglavnom izgrađeni od spirala, i dijelovi, sastavljeni od anti-paralelnih kuglica Inzulin

 4. / -Proteini l l -Spirale i -strukture nastaju duž toka Lancuga. Većina -struktura (paralelnih) je lokalizirana u središnjem dijelu molekule, gdje se te strukture savijaju pri izgledu propelera (“ twist” struktura), stvarajući krutu “bazu””, za koju su povezani drugi dijelovi molekule
4. / -Proteini l l -Spirale i -strukture nastaju duž toka Lancuga. Većina -struktura (paralelnih) je lokalizirana u središnjem dijelu molekule, gdje se te strukture savijaju pri izgledu propelera (“ twist” struktura), stvarajući krutu “bazu””, za koju su povezani drugi dijelovi molekule


 Domene l l l Prosječna veličina domene je 100 – 150 dodatnih, što označava kuglice promjera približno 2,5 nm. e. formiranje tercijarne strukture, koje se događa u završnim fazama formiranja globula
Domene l l l Prosječna veličina domene je 100 – 150 dodatnih, što označava kuglice promjera približno 2,5 nm. e. formiranje tercijarne strukture, koje se događa u završnim fazama formiranja globula

 Tercijarna struktura proteina... l l l karakterizira široku distribuciju uređenih i amorfnih dijelova u polipeptidnoj lanceti kao cjelini, što je posljedica interakcije bioloških radikala i leži u njihovoj vrsti i konformaciji. Dakle, tercijarna struktura opisuje prostorni raspored svih proteinskih molekula, budući da ih čini jedna polipeptidna tvar izravno povezana s oblikom proteinskih molekula, koji može varirati od končastog do končastog
Tercijarna struktura proteina... l l l karakterizira široku distribuciju uređenih i amorfnih dijelova u polipeptidnoj lanceti kao cjelini, što je posljedica interakcije bioloških radikala i leži u njihovoj vrsti i konformaciji. Dakle, tercijarna struktura opisuje prostorni raspored svih proteinskih molekula, budući da ih čini jedna polipeptidna tvar izravno povezana s oblikom proteinskih molekula, koji može varirati od končastog do končastog
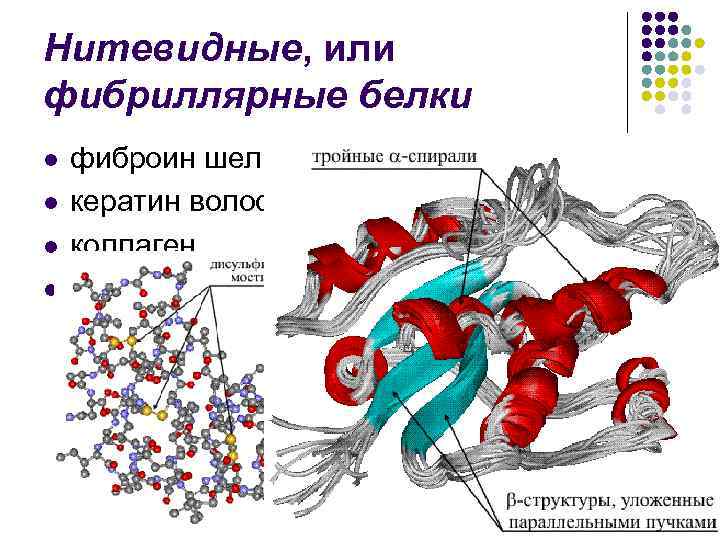 Nitasti ili fibrilarni proteini l l fibroin spaja keratin kose, rogova, akumulira kolagen i dr.
Nitasti ili fibrilarni proteini l l fibroin spaja keratin kose, rogova, akumulira kolagen i dr.

 Tercijarna struktura proteina temelj je funkcionalnosti proteina, što zahtijeva preciznu prostornu organizaciju velikih ansambala stvorenih u nedostatku aminokiselinskih viška. protein (denaturacija) mora se provoditi sve dok ne izgubi sposobnost funkcioniranja
Tercijarna struktura proteina temelj je funkcionalnosti proteina, što zahtijeva preciznu prostornu organizaciju velikih ansambala stvorenih u nedostatku aminokiselinskih viška. protein (denaturacija) mora se provoditi sve dok ne izgubi sposobnost funkcioniranja
 Tercijarna struktura proteina l l l Stabilnost tercijarne strukture leži u sustavu nekovalentnih interakcija u sredini proteinske globule (?) Neki proteini su dodatno stabilizirani kovalentnim - disulfidnim - vezama Glanzug = RTln. K = Nlanac – T S lanceug Nlanac 0 (S
Tercijarna struktura proteina l l l Stabilnost tercijarne strukture leži u sustavu nekovalentnih interakcija u sredini proteinske globule (?) Neki proteini su dodatno stabilizirani kovalentnim - disulfidnim - vezama Glanzug = RTln. K = Nlanac – T S lanceug Nlanac 0 (S
 Tercijarna struktura proteina l Spivvídnosti Chain i -T Schip leži u: l l broju nekovalentnih veza koje se postavljaju kada se proteinske molekule progutaju u globulu, a nakon polipeptidnog lancera ovisi o temperaturi. Zi zrostannyam T |T S| rano ili kasno krenuti | N| ta otvorena struktura gubi stabilnost – toplinska denaturacija proteina
Tercijarna struktura proteina l Spivvídnosti Chain i -T Schip leži u: l l broju nekovalentnih veza koje se postavljaju kada se proteinske molekule progutaju u globulu, a nakon polipeptidnog lancera ovisi o temperaturi. Zi zrostannyam T |T S| rano ili kasno krenuti | N| ta otvorena struktura gubi stabilnost – toplinska denaturacija proteina
 Tercijarna struktura proteina l l Kada se globula proguta, mijenja se entropija peptidnog koplja, ali istovremeno dolazi do povećanja entropije originatora - vode, što ima ključnu ulogu u stabilizaciji tercijarne strukture
Tercijarna struktura proteina l l Kada se globula proguta, mijenja se entropija peptidnog koplja, ali istovremeno dolazi do povećanja entropije originatora - vode, što ima ključnu ulogu u stabilizaciji tercijarne strukture
 Tercijarna struktura proteina l l Voda je nevažan, strukturni agens visokog reda (?) Polarne skupine proteina lako se ugrađuju u strukture vode poput leda, a vodene veze se umeću iz nje. Nepolarne skupine zauzimaju vlastite prazne prostore koje stvaraju molekule voda, vezana vodenim vezama Sve ovo isključuje kaotičan protok molekula vode, tako da postoji red, promjena entropije vode
Tercijarna struktura proteina l l Voda je nevažan, strukturni agens visokog reda (?) Polarne skupine proteina lako se ugrađuju u strukture vode poput leda, a vodene veze se umeću iz nje. Nepolarne skupine zauzimaju vlastite prazne prostore koje stvaraju molekule voda, vezana vodenim vezama Sve ovo isključuje kaotičan protok molekula vode, tako da postoji red, promjena entropije vode
 Tercijarna struktura proteina l l l Ako fragmenti nepolarnih skupina proteina ne mogu "sferom" izaći iz molekule, formira se globula u kojoj se značajan dio (ne manje od ½) hidrofobnih skupina pojavljuje u kontaktu s voda To podrazumijeva ugradnju hidrofobnih kontakata, van der Waalsovih kontakata. sile Dolazi do smanjenja entropije proteina. Nakon formiranja globule dolazi do propadanja značajnog dijela vodenih ligamenata i hidrofobnih interakcija proteinskih skupina s molekulama vode. Kaos ostatka se povećava, a povećava se entropija vode
Tercijarna struktura proteina l l l Ako fragmenti nepolarnih skupina proteina ne mogu "sferom" izaći iz molekule, formira se globula u kojoj se značajan dio (ne manje od ½) hidrofobnih skupina pojavljuje u kontaktu s voda To podrazumijeva ugradnju hidrofobnih kontakata, van der Waalsovih kontakata. sile Dolazi do smanjenja entropije proteina. Nakon formiranja globule dolazi do propadanja značajnog dijela vodenih ligamenata i hidrofobnih interakcija proteinskih skupina s molekulama vode. Kaos ostatka se povećava, a povećava se entropija vode

 Kvartarna struktura proteina... l l l Kvarterna struktura se nalazi u oligomernim proteinima, čije se molekule sastoje od dva ili više polipeptidnih koplja povezanih nekovalentno. ansambl globula) djeluje kao jedna molekula, u isto vrijeme koža globula koje su kombinirane zadržava značajnu autonomiju.
Kvartarna struktura proteina... l l l Kvarterna struktura se nalazi u oligomernim proteinima, čije se molekule sastoje od dva ili više polipeptidnih koplja povezanih nekovalentno. ansambl globula) djeluje kao jedna molekula, u isto vrijeme koža globula koje su kombinirane zadržava značajnu autonomiju.

 Kvartarna struktura proteina l l Podjedinice koje stvaraju kvaternarnu strukturu proteina mogu se potpuno razlikovati od njihovih stvarnih funkcija - heteromernih proteina, što vam omogućuje kombiniranje niza međusobno ovisnih funkcija u jednoj strukturi, stvarajući podnu ifunkcionalnu molekulu l l App. Protein kinaza: C-podjedinica je odgovorna za enzimsku aktivnost, a R-podjedinica je regulatorna.U homomernim proteinima podjedinice su iste
Kvartarna struktura proteina l l Podjedinice koje stvaraju kvaternarnu strukturu proteina mogu se potpuno razlikovati od njihovih stvarnih funkcija - heteromernih proteina, što vam omogućuje kombiniranje niza međusobno ovisnih funkcija u jednoj strukturi, stvarajući podnu ifunkcionalnu molekulu l l App. Protein kinaza: C-podjedinica je odgovorna za enzimsku aktivnost, a R-podjedinica je regulatorna.U homomernim proteinima podjedinice su iste
 Kvartarna struktura proteina l Međupodjedinični kontakti – sustav nekovalentnih interakcija je već razvijen.Kvartarna struktura je manja, manje tercijarna, što znači da ima manje hidrofobnih kontakata.
Kvartarna struktura proteina l Međupodjedinični kontakti – sustav nekovalentnih interakcija je već razvijen.Kvartarna struktura je manja, manje tercijarna, što znači da ima manje hidrofobnih kontakata.

 Funkcije četvrtinske strukture 1. Kombinacija mnogih međusobno povezanih funkcija u jednu strukturu 2. Arhitektonska funkcija 1. Oblikovanje prostranih struktura u ravnomjerne sklopive konfiguracije kako bi se osigurao specifičan i funkcionalni kapacitet proteina (feritin) 2. Sažetak naknadnih enzimskih reakcija 3. Formiranje funkcionalnog
Funkcije četvrtinske strukture 1. Kombinacija mnogih međusobno povezanih funkcija u jednu strukturu 2. Arhitektonska funkcija 1. Oblikovanje prostranih struktura u ravnomjerne sklopive konfiguracije kako bi se osigurao specifičan i funkcionalni kapacitet proteina (feritin) 2. Sažetak naknadnih enzimskih reakcija 3. Formiranje funkcionalnog
 Funkcije četvrtinske strukture 3. Osiguravanje višestrukih interakcija proteina s proširenim strukturama DNA-vezujući proteini – dimeri (?) 4. Regulatorna funkcija. Prijenos učinka (oštećenja tercijarne strukture pri interakciji sa supstratom) prenosi se s jedne podjedinice na drugu, što dovodi do obnove svih kvarternih struktura.
Funkcije četvrtinske strukture 3. Osiguravanje višestrukih interakcija proteina s proširenim strukturama DNA-vezujući proteini – dimeri (?) 4. Regulatorna funkcija. Prijenos učinka (oštećenja tercijarne strukture pri interakciji sa supstratom) prenosi se s jedne podjedinice na drugu, što dovodi do obnove svih kvarternih struktura.


 1. Pojedinačni protein kože karakterizira jedinstvena struktura, koja osigurava jedinstvenost njegove funkcije 2. Stoga, razumijevanje strukture različitih proteina može biti ključ za razumijevanje prirode živih sustava i, stoga, očito, bitno
1. Pojedinačni protein kože karakterizira jedinstvena struktura, koja osigurava jedinstvenost njegove funkcije 2. Stoga, razumijevanje strukture različitih proteina može biti ključ za razumijevanje prirode živih sustava i, stoga, očito, bitno
 Literatura Berezov T. T., Korovkin B. F. Biološka kemija. - M.: Medicina, 1983 Metzler D. Biokemija. Kemijske reakcije u živim stanicama. U 3 sveska - T. svezak 1, 2. - M.: Svit, 1980 Ovchinnikov Yu.A. Bioorganska kemija. - M.: Prosvitnitstvo, 1987 Osnove biokemije / Ed. A. A. Anisimova. - M.: Vishcha Shkola, 1986 Rhys E., Strenberg M. Od stanica do atoma. Ilustrirani uvod u molekularnu biologiju. - M.: Svit, 1988 Stepanov V. M. Molekularna biologija. Struktura i funkcija proteina. - M.: Vishcha škola, 1996 Pilipovich Yu B. Osnove biokemije. - M.: Vishcha škola, 1993
Literatura Berezov T. T., Korovkin B. F. Biološka kemija. - M.: Medicina, 1983 Metzler D. Biokemija. Kemijske reakcije u živim stanicama. U 3 sveska - T. svezak 1, 2. - M.: Svit, 1980 Ovchinnikov Yu.A. Bioorganska kemija. - M.: Prosvitnitstvo, 1987 Osnove biokemije / Ed. A. A. Anisimova. - M.: Vishcha Shkola, 1986 Rhys E., Strenberg M. Od stanica do atoma. Ilustrirani uvod u molekularnu biologiju. - M.: Svit, 1988 Stepanov V. M. Molekularna biologija. Struktura i funkcija proteina. - M.: Vishcha škola, 1996 Pilipovich Yu B. Osnove biokemije. - M.: Vishcha škola, 1993


