يتم تحديد بنية بروتين واحد من خلال مجموعة من الجينات. البروتينات: بنية البروتين الأولية، مخطط تكوين ثلاثي الببتيد
بيلكي- مركبات عضوية عالية الجزيئات تتشكل من زيادة أحماض ألفا الأمينية.
ش مستودع البروتينتشمل الفحم، الماء، النيتروجين، كيسين، سيركا. تشكل بعض البروتينات معقدات مع جزيئات أخرى، مثل الفوسفور والحديد والزنك والنحاس.
البروتينات لها وزن جزيئي مرتفع: زلال البيض - 36000، الهيموجلوبين - 152000، الميوسين - 500000. للمقارنة: الوزن الجزيئي للكحول - 46، حمض الأوتيك - 60، البنزين - 78.
مستودع الأحماض الأمينية للبروتينات
بيلكي- البوليمرات غير الدورية مثل المونومرات الأحماض الأمينية ألفا. هناك 20 نوعًا من الأحماض الأمينية ألفا تسمى مونومرات البروتين، على الرغم من أنه تم تحديد أكثر من 170 نوعًا في الخلايا والأنسجة.
ومن المهم الإشارة إلى أن الأحماض الأمينية يمكن تصنيعها في جسم الإنسان والمخلوقات الأخرى، وهي تنقسم إلى: استبدال الأحماض الأمينية- يمكن توليفها. الأحماض الأمينية الأساسية- لا يمكنهم التوليف. توجد الأحماض الأمينية الأساسية في الجسم مباشرة من الجسم. تقوم النباتات بتصنيع جميع أنواع الأحماض الأمينية.
مخزنة في تخزين الأحماض الأمينية، البروتينات على قدم وساق- ضباب مجموعة كاملة من الأحماض الأمينية. السفلي- جميع الأحماض الأمينية متوفرة يومياً. وبما أن البروتينات تتكون من أحماض أمينية، فإنها تسمى أنا آسف. عندما تحتوي البروتينات، بالإضافة إلى الأحماض الأمينية، على مكون آخر من الأحماض غير الأمينية (المجموعة الاصطناعية)، فإنها تسمى قابلة للطي. يمكن تمثيل المجموعة الاصطناعية بالمعادن (البروتينات المعدنية)، والكربوهيدرات (البروتينات السكرية)، والدهون (البروتينات الدهنية)، والأحماض النووية (البروتينات النووية).
شارب الأحماض الأمينية تنتقم: 1) مجموعة الكربوكسيل (-COOH)؛ 2) المجموعة الأمينية (-NH 2)؛ 3) مجموعة جذرية أو R (جزيء ريشتا). تختلف جذور بودوفا في أنواع مختلفة من الأحماض الأمينية. اعتمادًا على عدد المجموعات الأمينية ومجموعات الكربوكسيل التي تدخل إلى مستودع الأحماض الأمينية، يتم تقسيمها إلى: الأحماض الأمينية المحايدةهناك مجموعة كربوكسيل واحدة ومجموعة أمينية واحدة؛ الأحماض الأمينية الأساسيةهناك أكثر من مجموعة أمينية؛ الأحماض الأمينية الحمضيةما هي أكثر من مجموعة الكربوكسيل؟
أحماض أمينية أشباه الموصلات الأمفوتيريةلذلك، في الغذاء، يمكن أن يكون سبب الرائحة الكريهة كل من الأحماض والقواعد. في أصناف المياه، تظهر الأحماض الأمينية في أشكال أيونية مختلفة.
رابط الببتيد
الببتيدي- المركبات العضوية التي تتكون من فائض من الأحماض الأمينية المرتبطة برابطة الببتيد.
يتبع إطلاق الببتيدات تفاعل تكثيف الأحماض الأمينية. عندما تتفاعل المجموعة الأمينية لأحد الأحماض الأمينية مع المجموعة الكربوكسيلية لحمض أميني آخر، تنشأ بينهما رابطة تساهمية من النيتروجين والكربوهيدرات، وهو ما يسمى الببتيد. من المهم فصل كمية الأحماض الأمينية الزائدة التي تدخل في مخزن الببتيد ثنائي الببتيدي، ثلاثي الببتيدي، رباعي الببتيديإلخ. يمكن تكرار تكوين الرابطة الببتيدية بعدة طرق. جلب هذا للضوء الببتيدات. في أحد طرفي الببتيد توجد مجموعة أمينية حرة (تسمى النهاية N)، وفي الطرف الآخر توجد مجموعة كربوكسيل حرة (تسمى النهاية C).

تنظيم بسيط لجزيئات البروتين
تعتمد الوظائف المحددة للبروتينات على التكوين المكاني لجزيئاتها، بالإضافة إلى ذلك، تقوم الخلايا بتشذيب البروتينات بشكل نشط وغير مرئي في شكل مفتوح، يشبه الرمح، لذلك يتم ملاحظة تشكيل وتورم البنية والتشكل التافهين. انظر 4 مناطق تنظيم واسع من البروتينات.
هيكل البروتين الأساسي- تسلسل ذوبان فائض الأحماض الأمينية من المشرط متعدد الببتيد لتكوين جزيء البروتين. الرابطة بين الأحماض الأمينية هي الببتيد.

نظرًا لأن جزيء البروتين يتكون من 10 فائض من الأحماض الأمينية فقط، فإن المتغيرات الممكنة نظريًا لجزيئات البروتين والتي تختلف في ترتيب إضافة الأحماض الأمينية هي 10 20 . مع 20 حمضًا أمينيًا، يمكنك دمجها في مجموعات أكثر اختلافًا. تم التعرف على حوالي عشرة آلاف بروتين مختلف في الإنسان، تختلف من نوع إلى آخر، وكذلك من بروتينات الكائنات الحية الأخرى.
يشير الهيكل الأساسي لجزيء البروتين إلى قوة جزيئات البروتين وتكوينها الواسع. يؤدي استبدال حمض أميني بآخر في سلسلة البولي ببتيد إلى تغيير في قوى البروتين ووظائفه. على سبيل المثال، يؤدي استبدال الحمض الأميني الجلوتاميك في الوحدة الفرعية للهيموجلوبين بفالين إلى حقيقة أن جزيء الهيموجلوبين ككل لا يمكن أن يفقد وظيفته الرئيسية - النقل الحمضي؛ في مثل هذه الحالات، يصاب الأشخاص بالمرض - فقر الدم المنجلي.
الهيكل الثانوي- يتم ترتيب الحنجرة في مشرط متعدد الببتيد في شكل حلزوني (يبدو وكأنه زنبرك ممتد). تتكون الملفات الحلزونية من روابط مائية تتشابك بين مجموعات الكربوكسيل ومجموعات الأمينو. تشارك جميع مجموعات CO وNH تقريبًا في إنشاء وصلات المياه. والرائحة الكريهة أضعف من رائحة الببتيد، ولكنها عند تكرارها تكون غنية بالغازات، مما يعطي هذه التغيرات الثبات والقسوة. على مستوى البنية الثانوية توجد البروتينات: الفيبروين (الخيط، الحرير)، الكيراتين (الشعر، الأظافر)، الكولاجين (الوتر).



هيكل تريتين- وقحة Lancyugiv نصف المحبوبة في Globuli، وviniki veneolikenny من vinikenni من Zv'yazkiv التسعين (vodnevikh، disulfīdni) هو إلحاق ميرا girdrophic المتبادلة بواسطة جذور الأحماض الأمينية. يتم لعب الدور الرئيسي للهيكل الثالث المضيء من خلال التفاعلات المحبة للماء. في الجزيئات المائية، تميل الجذور الكارهة للماء إلى التقارب في الماء، وتتجمع في منتصف الكرية، بينما تميل الجذور المحبة للماء، نتيجة للترطيب (التفاعل مع ثنائيات أقطاب الماء)، إلى الاستقرار على سطح الجزيء. في بعض البروتينات، يتم تثبيت البنية الثلاثية عن طريق الروابط التساهمية ثاني كبريتيد التي تتشكل بين ذرات الكربون لاثنين من السيستين الزائدة. على مستوى البنية الثلاثية توجد إنزيمات وأجسام مضادة وهرمونات معينة.

هيكل رباعيوفي البروتينات القابلة للطي، الجزيئات التي تتشكل إلى كرتين أو أكثر. يتم دمج الوحدات الفرعية في الجزيء من خلال التفاعلات الأيونية والكارهة للماء والكهروستاتيكية. في بعض الأحيان، عندما يتم تشكيل البنية الرباعية، تظهر روابط ثاني كبريتيد بين الوحدات الفرعية. أكبر بروتين له بنية ربعية هو الهيموجلوبين. يحتوي على وحدتين فرعيتين α (141 فائض من الأحماض الأمينية) ووحدتين فرعيتين β (146 فائض من الأحماض الأمينية). يرتبط جزيء الهيم بوحدة فرعية من الجلد، وهي المسؤولة عن إطلاقه.
إذا انحرف تكوين البروتين عن الطبيعي لأي سبب من الأسباب، فلن يتمكن البروتين من الاحتفاظ بوظائفه. على سبيل المثال، سبب "قصة البقرة" (اعتلال الدماغ الإسفنجي) هو التشكل غير الطبيعي للبريونات - البروتينات السطحية للخلايا العصبية.
قوة السناجب
تخزين الأحماض الأمينية، وهيكل جزيء البروتين يعني قوة. تكتسب البروتينات حموضتها الأساسية، والتي تحددها جذور الأحماض الأمينية: كلما زادت حمضية البروتين، كلما كان التعبير عن حموضته أكثر سطوعًا. قيمة العطاء والقدوم H+ تعني القوة العازلة للبروتينات; أحد أهم الحواجز المؤقتة هو الهيموجلوبين في خلايا الدم الحمراء، والذي يحافظ على درجة حموضة الدم عند مستوى ثابت. - بروتينات منفصلة (الفبرينوجين)، وبروتينات غير منفصلة تساهم في الوظائف الميكانيكية (الفبرين، الكيراتين، الكولاجين). هناك بروتينات نشطة في شكل كيميائي (إنزيمات)، وهناك بروتينات غير نشطة كيميائيا، مقاومة لتدفق العقول المختلفة من العالم الخارجي وغير مستقرة للغاية.
العوامل الخارجية (التدفئة، التعرض للأشعة فوق البنفسجية، المعادن المهمة وأملاحها، تغيرات الرقم الهيدروجيني، الإشعاع، تلوث المياه)
قد يسبب ضررا للتنظيم الهيكلي لجزيء البروتين. تسمى عملية فقدان التشكل التافه، وهو الشكل السائد لجزيء البروتين تمسخ. سبب تمسخ الطبيعة هو تمزق الأربطة التي تعمل على تثبيت بنية البروتين. إن أضعف الأربطة في الكلى تتمزق، وعندما تكون العقول أقوى تمزق الأقوياء. ثم يتم تطوير الهياكل الرباعية ثم الثلاثية والثانوية. يؤدي تغيير التكوين المكاني إلى تغيير في قوة البروتين، ونتيجة لذلك، يجعل من المستحيل على البروتين أن يفقد السيطرة على وظائفه البيولوجية. وبما أن تمسخ الطبيعة لا يصاحبه تدمير البنية الأولية، فإنه يجوز ذلك بالذئب، بمجرد حدوث تجديد ذاتي لتشكل بروتين الطاقة. يتم التعرف على هذا التمسخ، على سبيل المثال، عن طريق بروتينات مستقبلات الغشاء. تسمى عملية تحديث بنية البروتين بعد تمسخه إعادة الطبيعة. إذا كان من المستحيل تجديد التكوين المكاني للبروتين، فسيتم استدعاء تمسخ البروتين غير قابل للإلغاء.
وظائف البروتينات
| وظيفة | يرجى تقديم تفسير |
|---|---|
| بوديفيلنا | تشارك البروتينات في تكوين الهياكل الخلوية وما بعد السريرية: فهي تدخل الأغشية الخلوية (البروتينات الدهنية والبروتينات السكرية) والشعر (الكيراتين) والأوتار (الكولاجين) وما إلى ذلك. |
| النقل | يضيف بروتين الهيموجلوبين في الدم الحموضة وينقلها من الدم إلى جميع الأنسجة والأعضاء، ومنها ينقل ثاني أكسيد الكربون؛ يشتمل تخزين أغشية الخلايا على بروتينات خاصة تضمن النقل النشط والانتقائي الصارم لبعض المواد والأيونات من الخلية إلى القلب الخارجي والعودة. |
| تنظيمية | تشارك هرمونات البروتين في تنظيم عمليات استقلاب الكلام. على سبيل المثال، ينظم هرمون الأنسولين مستوى الجلوكوز في الدم، ويعزز تخليق الجليكوجين، ويزيد من تقليل الدهون من الكربوهيدرات. |
| زهيسنا | عندما تخترق البروتينات والكائنات الحية الدقيقة الأجنبية (المستضدات) الجسم، يتم إنشاء بروتينات خاصة - الأجسام المضادة، التي تربطها وتقشرها. الفيبرين، الذي يتم إنشاؤه مع الفيبرينوجين، يقلل النزيف. |
| دفيجونا | تضمن بروتينات الأكتين والميوسين قصيرة العمر تقصير مدة القرحة في الحيوانات الغنية بالخلايا. |
| سيجنالنا | يوجد عند الغشاء السطحي للخلية جزيئات بروتينية تغير بنيتها الثلاثية استجابة لعوامل الوسط الخارجي، وبالتالي تستقبل الإشارات من الوسط الخارجي وتنقل الأوامر إلى الخلية. |
| تخزين | في جسم الحيوانات، كقاعدة عامة، لا يتم تخزين البروتينات، الجناة: زلال البيض، كازين الحليب. ومع ذلك، يمكن تزويد البروتينات الموجودة في الجسم باحتياطي من كمية معينة من الكلام، على سبيل المثال، عندما ينهار الهيموجلوبين، لا يُفرز السائل من الجسم، بل يتم تخزينه، مما يخلق مركبًا يحتوي على بروتين الفيريتين. |
| نشيط | عندما يتحلل 1 جرام من البروتين إلى المنتجات النهائية، يظهر 176 كيلوجول. في البداية، تتحلل البروتينات إلى أحماض أمينية، ثم إلى المنتجات النهائية – الماء وثاني أكسيد الكربون والأمونيا. ومع ذلك، كمصدر للطاقة، لا يتم امتصاص البروتينات إلا إذا تم استهلاك مصادر أخرى (الكربوهيدرات والدهون). |
| المحفز | من أهم وظائف البروتينات. مزود بالبروتينات - الإنزيمات التي تعمل على تسريع التفاعلات الكيميائية الحيوية التي تحدث في الخلايا. على سبيل المثال، يحفز كربوكسيلاز ثنائي فوسفات الريبولوز التثبيت 2 أثناء عملية التمثيل الضوئي. |
تخمير
تخمير، أو الانزيمات- فئة خاصة من البروتينات التي تعتبر محفزات بيولوجية. بسبب الإنزيمات، تتم التفاعلات الكيميائية الحيوية بميوعة كبيرة. ترجع سرعة التفاعلات الأنزيمية بعشرات الآلاف من المرات (وبملايين المرات) إلى سرعة التفاعلات التي تحدث بمشاركة المحفزات غير العضوية. يسمى Rechovina، وهو ما يعطي الإنزيم تأثيره المادة المتفاعلة.
الإنزيمات - البروتينات الكروية، خصوصيات بودوفييمكن تقسيم الإنزيمات إلى مجموعتين: بسيطة ومعقدة. أنا آسف التخمرمع أبسط البروتينات، إذن. تتشكل في المقام الأول من الأحماض الأمينية. الانزيمات القابلة للطيє طي البيض ثم. قبل مستودعاتهم، بالإضافة إلى الجزء البروتيني، هناك مجموعة من الطبيعة غير البروتينية. العامل المساعد. في بعض الإنزيمات، تعمل الفيتامينات كعوامل مساعدة. يحتوي جزيء الإنزيم على جزء خاص يسمى المركز النشط. مركز نشط- جزء صغير من الإنزيم (من ثلاثة إلى اثني عشر فائضًا من الأحماض الأمينية)، والذي يربط الركيزة أو الركائز بمركب الإنزيم-الركيزة. بعد الانتهاء من التفاعل، يتحلل مركب الإنزيم والركيزة إلى الإنزيم ومنتج (منتجات) التفاعل. الانزيمات الفعلية نشطة (الكريم النشط) مراكز تفارغي- الأقسام التي تستخدم فيها منظمات السيولة والإنزيمات ( الانزيمات allosteric).

تتميز تفاعلات الحفز الأنزيمي بما يلي: 1) الكفاءة العالية، 2) الانتقائية الصارمة ومباشرة العمل، 3) خصوصية الركيزة، 4) التنظيم الدقيق والدقيق. يتم تفسير خصوصية الركيزة والتفاعل لتفاعلات الحفز الأنزيمي من خلال فرضيات E. Fisher (1890) وD. Koshland (1959).
هـ. فيشر (فرضية قفل المفتاح)على افتراض أن التكوينات الواسعة للمركز النشط للإنزيم والركيزة هي نفسها تمامًا. الركيزة هي نفس "المفتاح"، والإنزيم هو "القفل".
د. كوشلاند (فرضية القفاز اليدوي)على افتراض أن التشابه في البنية بين الركيزة والمركز النشط للإنزيم يتم إنشاؤه فقط في لحظة تفاعلهما واحدًا تلو الآخر. وتسمى هذه الفرضية أيضا فرضية الأنواع المستحثة.
سيولة التفاعلات الأنزيمية تعتمد على: 1) درجة الحرارة؛ 2) تركيز الانزيم. 3) تركيز الركيزة. 4) الرقم الهيدروجيني. تجدر الإشارة إلى أن شظايا الإنزيم هي بروتينات، فإن نشاطها هو الأعلى في العقول الطبيعية من الناحية الفسيولوجية.
لا يمكن معالجة معظم الإنزيمات إلا عند درجات حرارة تتراوح من 0 إلى 40 درجة مئوية. في هذه المناطق، تزداد سيولة التفاعل مرتين تقريبًا عندما ترتفع درجة حرارة الجلد إلى 10 درجات مئوية. عند درجات حرارة أعلى من 40 درجة مئوية، يخضع البروتين لعملية تمسخ ويتناقص نشاط الإنزيم. عند درجات حرارة قريبة من نقطة التجمد، يتم تعطيل الإنزيمات.
مع زيادة كمية الركيزة، تزداد سيولة التفاعل الأنزيمي حتى يصبح عدد الجزيئات في الركيزة مساوياً لعدد الجزيئات في الإنزيم. مع زيادة أخرى في كمية الركيزة، لا تزيد السيولة، طالما أن المراكز النشطة للإنزيم تصبح مشبعة. تؤدي التركيزات الأعلى من الإنزيم إلى زيادة النشاط التحفيزي، لأنه خلال ساعة واحدة من التفاعل يتعرض عدد أكبر من الجزيئات للركيزة.

بالنسبة لإنزيم الجلد، هناك قيمة pH مثالية يظهر عندها أقصى نشاط (الببسين – 2.0، الأميليز المخاطي – 6.8، الليباز تحت الجلد – 9.0). عند قيم pH أعلى أو أقل، ينخفض نشاط الإنزيم. في حالة التغيرات الشديدة في الرقم الهيدروجيني، يتم تغيير طبيعة الإنزيم.
يتم تنظيم سيولة الإنزيمات التفارغية عن طريق المواد التي تصل إلى مراكز التفارغ. ولأن هذه الكلمات تسرع من رد الفعل، فإنها تسمى نتنة المنشطات، ياكشو إلى جالم مثبطات.
تصنيف الانزيمات
بناءً على نوع التفاعلات الكيميائية المحفزة، تنقسم الإنزيمات إلى 6 فئات:
- إنزيم الأكسيدوكتاز(نقل الذرات من الماء أو الحمض أو الإلكترونات من مادة إلى أخرى - نازعة الهيدروجين)،
- ترانسفيراسي(نقل مجموعات الميثيل أو الأسيل أو الفوسفات أو الأمينية من جزيء إلى آخر - الترانساميناسات)،
- هيدرولاز(تفاعلات التحلل المائي، حيث يتم تصنيع منتجين من الركيزة - الأميليز، الليباز)،
- com.liazi(إضافة غير مائي إلى الركيزة أو الانفصال عن مجموعة جديدة من الذرات، حيث يمكن أن تنكسر الروابط CC، C-N، CO، C-S decarboxylase)،
- إيزوميراز(بيريبودوفا داخل الجزيئات - إيزوميراز)،
- ligazi(تكوين جزيئين نتيجة تكوين الروابط CC، C-N، CO-O، CS-synthetase).
تصنيف الرسومات الخاصة بك إلى فئات فرعية وفئات فرعية. وفي التصنيف الدولي الموجود فإن إنزيم الجلد له كود فريد يتكون من أربعة أرقام مفصولة بنقاط. الرقم الأول هو فئة، والآخر فئة فرعية، والثالث هو فئة فرعية، والرابع هو الرقم التسلسلي للإنزيم في تلك الفئة الفرعية، على سبيل المثال، رمز أرجيناز هو 3.5.3.1.
اذهب إلى المحاضرات رقم 2"ما هي وظائف الكربوهيدرات والدهون"
اذهب إلى المحاضرات رقم 4"وظائف الأحماض النووية ATP"
التخليق الحيوي للبروتين.
1. يتم تحديد بنية بروتين واحد:
1) مجموعة من الجينات 2) جينوم واحد
3) جزيء DNA واحد. 4) مجموع الجينات في الكائن الحي
2. يشفر الجين معلومات حول تسلسل المونومرات في الجزيء:
1) الحمض الريبي النووي النقال 2) AA 3) الجليكوجين 4) الحمض النووي
3. تسمى الثلاثيات بالمضادات:
1) DNA 2) t-RNA 3) i-RNA 4) r-RNA
4. يتطور التبادل البلاستيكي بشكل مهم مع التفاعل:
1) تفكك الخطب العضوية. 2) تفكك الخطب غير العضوية
3) تركيب المواد العضوية. 4) تركيب المواد غير العضوية
5. يحدث تخليق البروتين في الخلايا بدائية النواة:
1) على الريبوسومات في النواة 2) على الريبوسومات في السيتوبلازم 3) في موقع الخلية
4) على السطح الخارجي للغشاء السيتوبلازمي
6. تبدأ عملية البث:
1) في السيتوبلازم 2) في النواة 3) في الميتوكوندريا
4) على أغشية الغشاء الإندوبلازمي القصير
7. يحدث التوليف على أغشية الغشاء الإندوبلازمي الحبيبي:
1) اعبي التنس المحترفين. 2) في الكربوهيدرات. 3) الدهون. 4) البروتينات.
8. رموز ثلاثية واحدة:
1. واحد AK 2 واحد علامة الجسم 3. جبيرة AK
9. اكتمل تخليق البروتين في الوقت الحالي
1. التعرف على الكودون بواسطة الكودون المضاد 2. ظهور "علامة تقسيم" على الريبوسوم
3. نقل i-RNA إلى الريبوسوم
10. العملية التي يتم من خلالها قراءة المعلومات من جزيئات الحمض النووي.
1. الترجمة 2. النسخ 3. التحويل
11. يتم تحديد قوة البروتينات...
1. البنية الثانوية للبروتين 2. البنية الأولية للبروتين
3. بنية البروتين الثلاثي
12. العملية التي يتم من خلالها التعرف على الكودون المضاد باعتباره كودونًا على i-RNA
13. مراحل التخليق الحيوي للبروتين.
1. النسخ، الترجمة 2. التحويل، الترجمة
3.التنظيم والنسخ
14. يتكون Anticodon t-RNA من نيوكليوتيدات UCG. ما هو الحمض النووي الثلاثي المكمل لyoumu؟
1.UUG 2.TTC 3.TCG
15. عدد t-RNAs الذي يشارك في الترجمة تقليدي:
1. كودونات i-RNA التي تشفر الأحماض الأمينية 2. جزيئات i-RNA
3 الجينات التي تتواجد قبل جزيء DNA 4. البروتينات التي يتم تصنيعها على الريبوسومات
16. تحديد تسلسل نيوكليوتيدات i-RNA أثناء النسخ من أحد فروع الحمض النووي: A-G-T-C-G
1) يو 2) ز 3) ج 4) أ 5) ج
17. أثناء تضاعف جزيء DNA، يتم إنشاء ما يلي:
1) الخيط الذي انقسم إلى أجزاء من جزيئات ابنة
2) جزيء يتكون من شريطين جديدين من الحمض النووي
3) جزيء نصفه يتكون من خيوط iRNA
4) جزيء ابنة يتكون من شريط DNA القديم والجديد
18. قالب تخليق جزيء iRNA أثناء النسخ هو:
1) جزيء DNA بأكمله 2) سطح أحد جزيئات DNA
3) قطعة من الحمض النووي Lancs
4) في بعض الأنواع، واحد من جزيء DNA، وفي أنواع أخرى - جزيء DNA بأكمله.
19. عملية الانغماس الذاتي لجزيء DNA.
1. النسخ المتماثل 2. الإصلاح
3. التناسخ
20. أثناء عملية التخليق الحيوي للبروتين في الخلايا، تكون طاقة ATP:
1) الإنفاق 2) المخزون
3) لم ينفق ولم يرى
21. في الخلايا الجسدية لكائن غني بالخلايا:
1) مجموعة مختلفة من الجينات والبروتينات 2) مجموعة جديدة من الجينات والبروتينات
3) مجموعة جديدة من الجينات، أو مجموعة مختلفة من البروتينات
4) مجموعة جديدة من البروتينات، أو مجموعة مختلفة من الجينات
22. واحد ثلاثي من الحمض النووي يحمل معلومات عن:
1) تسلسل الأحماض الأمينية في جزيء البروتين
2) علامات الجسم 3) الأحماض الأمينية في جزيء البروتين المركب
4) تخزين جزيء الحمض النووي الريبي (RNA).
23. أي من العمليات غير موجودة في خلايا أي وظيفة:
1) تخليق البروتين 2) استقلاب البروتين 3) الانقسام 4) الانقسام الاختزالي
24. يشير مفهوم "النسخ" إلى العملية:
1) اندساس الحمض النووي 2) تخليق الحمض النووي الريبي (i-RNA) على الحمض النووي
3) انتقال i-RNA إلى الريبوسومات 4) تكوين جزيئات البروتين على الجسيمات المتعددة
25. جزء من جزيء DNA يحمل معلومات حول جزيء بروتين واحد:
1) الجين 2) النمط الظاهري 3) الجينوم 4) النمط الجيني
26. يحدث النسخ في حقيقيات النوى في:
1) السيتوبلازم 2) الغشاء الإندوبلازمي 3) الليزوزومات 4) النوى
27. يتم الحصول على تخليق البروتين من :
1) الشبكة الإندوبلازمية الحبيبية
2) الشبكة الإندوبلازمية الملساء 3) النواة 4) الليزوزومات
28. يتم ترميز حمض أميني واحد:
1) عدة نيوكليوتيدات 2) نيوكليوتيدتان
3) نيوكليوتيدات واحدة. 4) ثلاث نيوكليوتيدات
29. يحتوي ثلاثي نيوكليوتيدات ATC في جزيء DNA على كودون مماثل لجزيء i-RNA:
1) العلامة 2) UAG 3) UTC 4) TsAU
30. علامات التقسيمالكود الجيني:
1. تشفير بروتينات الأغنية 2. تحفيز تخليق البروتين
3. تعزيز تخليق البروتين
31. عملية الاشتراك الذاتي لجزيء الحمض النووي.
1. النسخ المتماثل 2. الإصلاح 3. إعادة الدمج
32. وظيفة i-RNA في عملية التخليق الحيوي.
1. حفظ معلومات الضغط 2. نقل AK إلى الريبوسومات
3. إمداد الريبوسومات بالمعلومات
33. العملية التي يقوم فيها t-RNA بجلب الأحماض الأمينية إلى الريبوسوم.
1. النسخ 2. الترجمة 3. التحول
34. الريبوسومات، التي تصنع نفس جزيء البروتين.
1. الكروموسوم 2. بوليزوم 3. ميجا كروموسوم
35. العملية التي تعمل من خلالها الأحماض الأمينية على تثبيت جزيء البروتين.
1. النسخ 2. الترجمة 3. التحول
36. قبل رد فعل تخليق المصفوفة، نأتي...
1. تكرار الحمض النووي 2. النسخ والترجمة 3. النوعان صحيحان
37. يحمل أحد توائم الحمض النووي معلومات حول:
1. تسلسل الأحماض الأمينية في جزيء البروتين
2. مكان غناء AK في مشرط البروتين
3. علامة كائن معين
4. الأحماض الأمينية التي يحتوي عليها بروتين لانسيت
38. قامت Genya بتشفير معلومات حول:
1) بنية البروتينات والدهون والكربوهيدرات 2) التركيب الأساسي للبروتين
3) تسلسل النيوكليوتيدات في الحمض النووي
4) تسلسل الأحماض الأمينية في 2 أو أكثر من جزيئات البروتين
39. يبدأ تركيب mRNA بما يلي:
1) انقسام الحمض النووي إلى شريطين. 2) التفاعل مع إنزيم بوليميراز RNA والجين
3) الجيل الفرعي للجين 4) تفكك الجين إلى نيوكليوتيدات
40. يتم توفير النسخ:
1) في النواة 2) في الريبوسومات 3) في السيتوبلازم 4) في قنوات EPS الملساء
41. لا يحدث تخليق البروتين على الريبوسومات في:
1) مرض السل الزبودنيك 2) البجولي 3) ذبابة الغاريق 4) العاثيات
42. أثناء الترجمة، تكون المصفوفة لتجميع البروتين متعدد الببتيد هي:
1) أحد سلاسل DNA. 2) أحد جزيء DNA
3) جزيء iRNA 4) في بعض الأشكال أحد خيوط الحمض النووي، وفي حالات أخرى - جزيء iRNA
البنية الأساسية للبروتينات عبارة عن رمح متعدد الببتيد خطي مع أحماض أمينية متصلة بواسطة روابط الببتيد. البنية الأولية هي أبسط مستوى من التنظيم الهيكلي لجزيء البروتين. يتم ضمان الثبات العالي من خلال روابط الببتيد التساهمية بين مجموعة α-amino لحمض أميني واحد ومجموعة α-carboxyl لحمض أميني آخر.
إذا شاركت المجموعة الأمينية من البرولين أو الهيدروكسي برولين، من حيث الرابطة الببتيدية، فسيبدو الأمر مختلفًا

عندما تتشكل الروابط الببتيدية في الكلى، يتم تنشيط مجموعة الكربوكسيل لأحد الأحماض الأمينية، ثم تتحد مع مجموعة أمينية أخرى. ومن الممكن أيضًا إجراء التوليف المختبري للبولي ببتيدات.
الارتباط الببتيد هو جزء من رمح متعدد الببتيد يكرر نفسه. هناك القليل من الخصائص التي لا تتعلق فقط بشكل البنية الأساسية، ولكن أيضًا بالمستوى الأكثر أهمية لتنظيم المشرط متعدد الببتيد:


· المستوى المشترك - جميع الذرات الموجودة في مجموعة الببتيد تقع في نفس المستوى.
· الصلاحية موجودة في شكلين رنينيين (شكل الكيتو والإينول)؛
· تبديل موضع الشفعاء في اتصال CN-N بنسبة مائة بالمائة؛
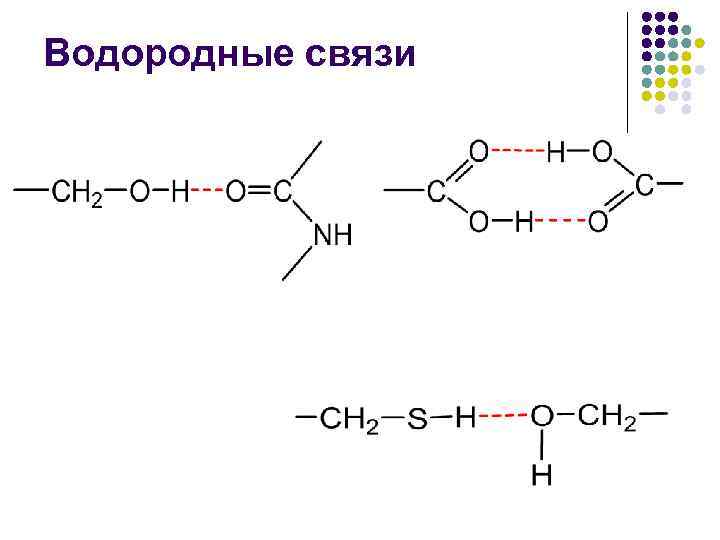
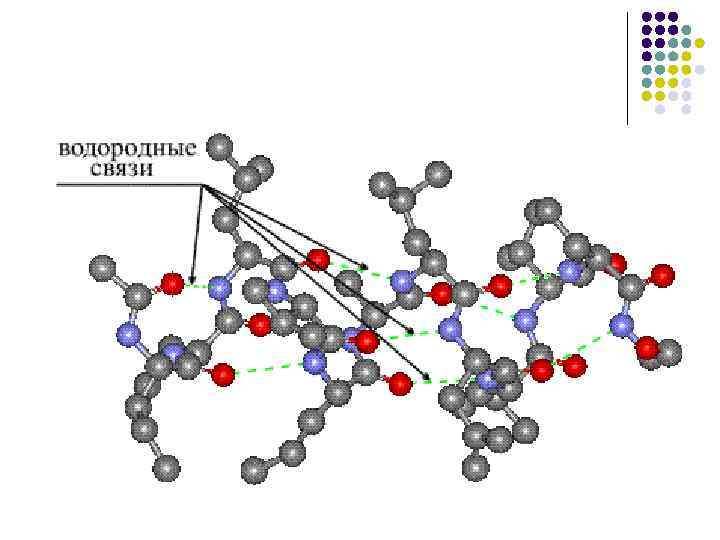
· وجود تكوين الأربطة المائية، حيث يمكن للجلد مع مجموعات الببتيد تكوين رباطين مائيين مع مجموعات أخرى، بما في ذلك المجموعات الببتيدية.
ترتبط مجموعات الببتيد بالمجموعة الأمينية من البرولين أو الهيدروكسي برولين. رائحة المبنى الكريهة تنشأ بصوت مائي واحد فقط (شيء رائع). يشار إلى ذلك على البنية الثانوية المصبوبة للبروتين. يتم تدمير مركب البولي ببتيد، الذي يحتوي على البرولين أو الهيدروكسي برولين، بسهولة ولا يمكن إزالته، كما هو الحال دائمًا، بواسطة الروابط المائية الأخرى.
مخطط صياغة ثلاثي الببتيد:

مستويات التنظيم المكاني للبروتينات: البنية الثانوية للبروتينات: مفاهيم الحلزون ألفا والكرة المطوية. البنية الثلاثية للبروتينات: مفاهيم حول البروتين الأصلي وتمسخ البروتين. يشبه التركيب الرباعي للبروتينات الهيموجلوبين.
البنية الثانوية للبروتين.تحت البنية الثانوية للبروتين، نفهم الطريقة التي يتم بها ترتيب رمح البولي ببتيد في بنية منظمة. وفقًا للتكوين، تكون العناصر التالية للهيكل الثانوي مرئية: α -حلزوني β -كرة أجزاء المستودع.
نموذج بودوفي α-اللوالب, الذي يحمي كل قوة الرابطة الببتيدية، تم تحليله بواسطة L. Pauling وR. Corey (1949 - 1951).
إلى الطفل 3، أالرسم البياني الموضح α -اللوالب، والتي تعطي معلومات حول المعلمات الرئيسية لحرق مشرط متعدد الببتيد α - حلزوني بحيث تكون المنعطفات الحلزونية منتظمة، بحيث يكون التكوين الحلزوني متماثلًا لولبيًا (الشكل 3، ب). على منعطف جلدي α -الحلزونات لديها فائض من الأحماض الأمينية قدره 3.6. الوقوف بين المنعطفات أو تصبح حافة اللولب 0.54 نانومتر، حيث يصل المنعطف التالي إلى 26 درجة. تشكيل وتقليم α - يتكون التكوين الحلزوني من بنية الأربطة المائية التي يتم إنشاؤها بين مجموعات الببتيد في الجلد ن-اذهب ثا ( ص+3) فائض الأحماض الأمينية. ورغم أن طاقة الأربطة المائية قليلة إلا أن كميتها عالية لتنتج تأثيرا طاقيا كبيرا نتيجة لذلك α - التكوين الحلزوني لإكمال الحامل. لا تشارك الجذور البيولوجية لتجاوزات الأحماض الأمينية في المادة α -التكوين الحلزوني، وبالتالي فإن جميع الأحماض الأمينية الفائضة فيه α -اللوالب متساوية.
البروتينات الطبيعية أقل يمينًا α - اللوالب.
كرة مطوية β- عنصر آخر من عناصر الهيكل الثانوي. على صفحة الإدارة α -اللوالب β - الكرة المطوية لها شكل خطي، وليس شكل ستريجنيفويو (الشكل 4). يتم تحديد البنية الخطية من خلال أصل الروابط المائية بين مجموعات الببتيد التي تقف على أقسام مختلفة من رمح متعدد الببتيد. يبدو أن هذه القطع قريبة من سطح وصلة المياه بين - C = O وHN - المجموعات (0.272 نانومتر).

صغير 4. الرسم التوضيحي التخطيطي β
- كرة متكررة للمستودع (الأسهم تشير
 حول مشرط متعدد الببتيد)
حول مشرط متعدد الببتيد)
صغير 3. المخطط ( أ) ذلك النموذج ( ب) α -اللوالب
يعتبر الهيكل الثانوي للبروتين هو الهيكل الأساسي. يتم إنشاء فائض من الأحماض الأمينية بكميات مختلفة قبل تكوين الأربطة المائية، والتي يتم بعد ذلك سكبها في الخليط. α -اللوالب أو β -شارو. تشمل الأحماض الأمينية القابلة للذوبان في الحلزون ألانين، وحمض الجلوتاميك، والجلوتامين، والليوسين، والليسين، والميثيونين، والهستيدين. إذا تم تشكيل جزء البروتين في المقام الأول من الأحماض الأمينية الزائدة، فسيتم تشكيله في هذه المرحلة α -حلزوني. فالين، آيزوليوسين، ثريونين، تيروزين وفينيل ألانين يوفرون الحل β - كرات من البولي ببتيد. تظهر الهياكل المضطربة على قطع رمح البولي ببتيد، حيث تتركز بقايا الأحماض الأمينية مثل الجليسين، السيرين، حمض الأسبارتيك، الأسباراجين، البرولين.
في البروتينات الغنية، على الفور α -اللوالب، ذلك β -شاري. يختلف جزء من التكوين الحلزوني في البروتينات المختلفة. وبالتالي، فإن بروتين اللحم باراميوزين يكون حلزونيًا بنسبة 100٪ تقريبًا؛ نسبة عالية من التكوين الحلزوني في الميوجلوبين والهيموجلوبين (75%). ومع ذلك، في التربسين والريبونوكلياز، جزء كبير من لانجوج متعدد الببتيد يتناسب مع شاروفيت β -الهياكل. تنتفخ بروتينات الأنسجة الداعمة - الكيراتين (بروتينات الشعر)، والكولاجين (بروتينات الجلد والأوتار). β -تكوين الرماح عديد الببتيد.
هيكل البروتين الثالث.البنية الثلاثية للبروتين هي وسيلة لوضع رمح متعدد الببتيد في الفضاء. لكي يضيف البروتين قوة إلى قوته الوظيفية، يجب على مشرط متعدد الببتيد أن يحترق منفردًا في الفضاء، مكونًا بنية نشطة وظيفيًا. ويسمى هذا الهيكل محلي. بغض النظر عن العدد الكبير من الهياكل المكانية الممكنة نظريًا لمشرط متعدد الببتيد المحيط، يجب إحضار البروتين الحنجري إلى تكوين تكوين أصلي واحد.
تثبيت البنية الثلاثية للبروتين عن طريق التفاعلات التي تحدث بين الجذور البيولوجية لفائض الأحماض الأمينية في أقسام مختلفة من رمح البولي ببتيد. يمكن تقسيم هذه التفاعلات إلى نقاط قوة ونقاط ضعف.
تتضمن التفاعلات القوية روابط تساهمية بين ذرات الكبريت الموجودة في السيستين الزائد، والتي تقف عند أجزاء مختلفة من رمح البولي ببتيد. وبخلاف ذلك، تسمى هذه الروابط جسور ثاني كبريتيد؛ يمكن تمثيل تشكيل جسر ثاني كبريتيد على النحو التالي:
إلى جانب الروابط التساهمية، يتأثر التركيب الثلاثي لجزيء البروتين بالتفاعلات الضعيفة، والتي تنقسم بطبيعتها إلى قطبية وغير قطبية.
تتضمن التفاعلات القطبية اتصالات أيونية ومائية. يتم إنشاء التفاعلات الأيونية عند الاتصال بين مجموعات موجبة الشحنة من مخلفات جذور الليسين والأرجينين والهستيدين ومجموعات COOH سالبة الشحنة من أحماض الأسبارتيك والجلوتاميك. تحدث الاتصالات المائية بين المجموعات الوظيفية من الجذور البيولوجية لتجاوزات الأحماض الأمينية.
التفاعلات غير القطبية أو تفاعلات فان دير فالس بين جذور الكربوهيدرات في بقايا الأحماض الأمينية تمنع تكوينها جوهر مسعور (بقع دهنية) في منتصف الكرية البروتينية، لأن سوف تختفي جذور الكربوهيدرات عند ملامستها للماء. كلما زاد عدد الأحماض الأمينية غير القطبية التي يحتوي عليها البروتين، زاد دور روابط فان دير فال في بنيته الثلاثية الشكل.
تشير الروابط العددية بين جذور البروتين للأحماض الأمينية الزائدة إلى التكوين المكاني لجزيء البروتين (الشكل 5).

صغير 5. أنواع الأربطة التي تدعم البنية الثلاثية للبروتين:
أ- خليط ثاني كبريتيد. ب -الارتباط الأيوني ج، د -توصيلات المياه؛
د -سندات فان دير فالس
إن البنية الثلاثية للبروتين المأخوذ حديثًا فريدة من نوعها، تمامًا كما أن البنية الأولية فريدة من نوعها. فقط المساحة الصحيحة لوضع البروتين هي التي ستبقيه نشطًا. يؤدي التدمير الهائل للبنية الثالثة إلى تغير في مستويات البروتين وفقدان النشاط البيولوجي.
التركيب الرباعي للبروتين.تتكون البروتينات التي يزيد وزنها الجزيئي عن 100 كيلو دالتون 1 عادةً من عدة رماح متعددة الببتيد ذات وزن جزيئي صغير. الهيكل الذي يتكون من عدد كبير من رماح البولي ببتيد، التي تشغل موقعًا ثابتًا تمامًا، على ما يبدو واحدة من الأخرى، ونتيجة لذلك يكون للبروتين نشاط مختلف، يسمى البنية الرباعية للبروتين. البروتين الذي له بنية ربعية يسمى جزيءاو اخرى متعدد ، ومن الواضح أن مستودعات مشارط البولي ببتيد الخاصة بها الوحدات الفرعية او اخرى بروتوميرات . القوة المميزة للبروتينات ذات البنية الرباعية هي تلك التي ليس لوحدتها الفرعية أي نشاط بيولوجي.
يتم تحقيق استقرار البنية الرباعية للبروتين من خلال تفاعل التفاعلات القطبية بين جذور الأحماض الأمينية المترجمة على سطح الوحدات الفرعية. مثل هذه التفاعلات مهمة لتقليل الوحدات الفرعية للمجمع المنظم. تسمى أقسام الوحدات الفرعية التي تتفاعل بميدان الاتصال.
المثال الكلاسيكي للبروتين الذي يحتوي على بنية ربعية هو الهيموجلوبين. جزيء الهيموجلوبين ذو الوزن الجزيئي 68000. ويتكون من أربع وحدات فرعية من نوعين مختلفين. α і β / α -تتكون الوحدة الفرعية من 141 حمض أميني وهي أ β - عز 146. هيكل التعليم العالي α - І β -الوحدة الفرعية مماثلة لوزنها الجزيئي (17000 لذلك). وحدة فرعية من الجلد للانتقام من المجموعة الاصطناعية - الهيم . توجد شظايا الهيم أيضًا في بروتينات أخرى (السيتوكروم، الميوجلوبين)، والتي تتشكل بشكل أكبر، ولكننا نود أن نناقش بإيجاز بنية تلك البروتينات (الشكل 6). الهيم المجمع هو نظام دوري متحد المستوى مطوي يتكون من ذرة مركزية، والتي تخلق روابط تنسيق مع بعض البوليمر الزائد، متصلة بواسطة مواقع الميثان (= CH -). في الهيموجلوبين، يبدأ مستوى الأكسدة بالحدوث (2+).
عدة وحدات فرعية - اثنان α و اثنان β - اتحدوا في هيكل واحد بهذه الطريقة α -الوحدات الفرعية الاتصال فقط β -الوحدات الفرعية ككل (الشكل 7).

صغير 6. هيكل الهيموجلوبين الهيم

صغير 7. تمثيل تخطيطي للتركيب الرباعي للهيموجلوبين:
الحديد - الهيموجلوبين
كما يمكن أن نرى من الطفل 7، جزيء واحد من الهيموجلوبين قادر على نقل 4 جزيئات من الحمض. يصاحب كل من الارتباط وإطلاق الحموضة تغييرات توافقية في الهيكل α - І β -وحدة فرعية من الهيموجلوبين وانحلالها المتبادل في الجزيء الحلقي. هذه الحقيقة تؤكد أن البنية الربعية للبروتين لم تعد صلبة تمامًا.
معلومات مماثلة.
إحدى ميزات البروتينات هي تنظيمها الهيكلي المعقد. تحتوي جميع البروتينات على بنية أولية وثانوية وثلاثية، وتلك التي تحتوي على اثنين أو أكثر من PPCs لها بنية رباعية (QS).
بنية البروتين الأولية (PSB) – هذا هو ترتيب ترسيب (تسلسل) فائض الأحماض الأمينية في القدرة الشرائية.
ومع ذلك، بسبب وفرة تكوينها والأحماض الأمينية، يمكن أن تكون البروتينات من أنواع مختلفة. على سبيل المثال، من اثنين من الأحماض الأمينية يمكنك الجمع بين نوعين مختلفين من ثنائي الببتيد:
مع كون عدد الأحماض الأمينية أكثر من 20، يكون عدد التركيبات الممكنة أقل من 210 18. وإذا كنت تعلم أن الحمض الأميني للبشرة في PPC يمكن أن يصبح حساسًا أكثر من مرة، فمن المهم حماية عدد الخيارات الممكنة.
أهمية بنية البروتين الأولية (PPS).
يمكن حساب PSB من البروتينات للحصول على المساعدة فينيلثيوهيدانتوين
طريقة
. تعتمد هذه الطريقة على رد الفعل المتبادل فينيل إيزوثيوسيانات
(FITC) مع α-AA. ونتيجة لذلك، يتم إنشاء مجمع من علاقتين FITC-AK
.
على سبيل المثال، دعونا ننظر إلى الببتيد ![]() باستخدام هذه الطريقة، يتم استخدام PSB لتحديد تسلسل فائض الأحماض الأمينية.
باستخدام هذه الطريقة، يتم استخدام PSB لتحديد تسلسل فائض الأحماض الأمينية.
يتفاعل FITC مع الحمض الأميني الطرفي (أ). يجري إنشاء المجمع فتغ-أالذي يضاف إلى المزيج ويشير إلى فعالية الحمض الأميني. أ. على سبيل المثال، تسي - asn
![]() إلخ. تقوية وتحديد جميع الأحماض الأمينية الأخرى باستمرار. إنها عملية شاقة. ويستمر بروتين PSB متوسط الحجم لعدة أشهر.
إلخ. تقوية وتحديد جميع الأحماض الأمينية الأخرى باستمرار. إنها عملية شاقة. ويستمر بروتين PSB متوسط الحجم لعدة أشهر.
فك تشفير PSB له الأولوية سينجيرو(1953) وهو اكتشاف الأنسولين PSB (الحائز على جائزة نوبل). يتكون جزيء الأنسولين من 2 PPCs - A و B.

يتكون A-Lancet من 21 حمضًا أمينيًا، و30 حمضًا أمينيًا. وترتبط PPC ببعضها البعض عن طريق مواقع ثاني كبريتيد. يصل عدد البروتينات التي تم تحديد PSBs الخاصة بها اليوم إلى 1500. يمكن للتغييرات الطفيفة في البنية الأولية أن تغير بشكل كبير قوة البروتين. يوجد HbA في كريات الدم الحمراء لدى الأشخاص الأصحاء - عند استبداله في -lancer HbA، في الموضع السادس gluعلى الفتحةاللوم على المرض الخطير فقر الدم المنجليلأن أي أطفال يولدون بهذه الحالة الشاذة يموتون مبكرًا. من ناحية أخرى، هناك خيارات محتملة لتغيير PSB، والتي لم يتم الإشارة إليها من قبل أي سلطات فيزيائية كيميائية وبيولوجية. على سبيل المثال، يوجد HbC في المركز السادس من b-lance بدلاً من gluliz، وقد لا يتنافس HbC مع قوته على HbA، والأشخاص الذين يحملون مثل هذا Hb في كريات الدم الحمراء لديهم يتمتعون بصحة جيدة عمليًا.
استقرار PSBويتم تأمينه بشكل رئيسي عن طريق روابط الببتيد التساهمية الداخلية، وبعبارة أخرى، عن طريق روابط ثاني كبريتيد.
البنية الثانوية للبروتين (PSB).
يُظهر PPV للبروتينات مرونة كبيرة ويطور بنية واسعة أو مميزة التشكل. في البروتينات، هناك مستويان من هذا التشكل – هذا هو VSB والبنية الثلاثية (تب).
VSB – يتوافق هذا التكوين لـ PPC، وطريقة وضعه أو لفه في أي شكل، مع البرنامج الموجود في PPCس.ب.
هناك ثلاثة أنواع رئيسية من VSB:
1) -حلزوني;
2) ب-بناء(كرة أجزاء المخزون أو ورقة أجزاء قابلة للطي) ؛
3) كرة بلا قلق.
-حلزوني .
تم اقتراح هذا النموذج بواسطة V. Pauling. هذا هو الأفضل للبروتينات الكروية. بالنسبة لأي نظام، الحالة الأكثر استقرارًا هي تلك التي تمثل الحد الأدنى من الطاقة الحرة. بالنسبة للببتيدات، يحدث هذا الموقف عندما ترتبط مجموعات CO وNH ببعضها البعض بواسطة رابطة مائية ضعيفة. ش أ -اللوالب تتفاعل مجموعة NH من بقايا الحمض الأميني الأول مع مجموعة CO من الحمض الأميني الرابع. ونتيجة لذلك، فإن العمود الفقري الببتيد يخلق دوامة، حيث يقع فائض 3.6 AA على الجلد.
1 حلزوني (دورة واحدة) = 3.6 تيار متردد = 0.54 نانومتر، مقطع - 26 درجة
لوحظ التواء PPC خلف سهم السنة، بحيث يتحرك اللولب إلى اليمين. من خلال الجلد 5 دورات (18 AC؛ 2.7 نانومتر) يتم تكرار تكوين PPC.
استقرار VSBأولاً قبل الروابط المائية، وفي الآخر - الببتيد وثاني كبريتيد. الروابط المائية أضعف بمقدار 10-100 مرة من الروابط الكيميائية القياسية؛ ومع ذلك، فإن الكمية الكبيرة من الرائحة الكريهة ستضمن قسوة وضغط VSB. تكون Lancelets على شكل حرف R حلزونية، مشطوفة إلى درجة الرنين ومنتشرة على جوانب ومحاور مختلفة.
ب -بناء .
هذه أجزاء مطوية من مؤامرة PPC، والتي تشبه شكل ورقة مطوية مثل الأكورديون. يمكن أن تكون كرات PPC متوازية، حيث أن الهجوم يبدأ عند الطرف N أو C.
نظرًا لأن الرماح الضيقة للكرة موجهة مع الأطراف القريبة N-C وC-N، فإن الروائح الكريهة تسمى ضد التوازي.

 موازي
موازي
![]() ضد التوازي
ضد التوازي
يحدث تكوين روابط مائية، كما هو الحال في الحلزون A، بين مجموعات CO وNH.

 تعد دراسة التنظيم الهيكلي للبروتينات إحدى المشكلات الرئيسية في الكيمياء الحيوية الحديثة، ولها أهمية علمية وعملية مهمة لفهم التوسع الكبير في وظائف البروتينات. هل هناك العشرات أو المئات من الأحماض الأمينية في جزيء البروتين؟
تعد دراسة التنظيم الهيكلي للبروتينات إحدى المشكلات الرئيسية في الكيمياء الحيوية الحديثة، ولها أهمية علمية وعملية مهمة لفهم التوسع الكبير في وظائف البروتينات. هل هناك العشرات أو المئات من الأحماض الأمينية في جزيء البروتين؟
 بروتينات إميل فيشر هي عبارة عن عديدات ببتيدات مطوية، حيث ترتبط الأحماض الأمينية معًا بواسطة روابط الببتيد (R-CO-NH-R)، والتي تتشكل أثناء تفاعل مجموعات الكربوكسيل والمجموعات الأمينية من الأحماض الأمينية.
بروتينات إميل فيشر هي عبارة عن عديدات ببتيدات مطوية، حيث ترتبط الأحماض الأمينية معًا بواسطة روابط الببتيد (R-CO-NH-R)، والتي تتشكل أثناء تفاعل مجموعات الكربوكسيل والمجموعات الأمينية من الأحماض الأمينية.
 الدليل التجريبي للنظرية l l l تحتوي البروتينات الطبيعية على عدد صغير من المجموعات المعايرة – COOH و –NH 2. أثناء عملية التحلل المائي للبروتين، يتم إنشاء عدد متكافئ من المجموعات المعايرة – COOH و –NH 2 تحت تأثير البروتينات. في وجود إنزيمات معينة، يتم تقسيم البروتينات إلى أجزاء محددة بدقة (عديدات الببتيد). ويؤكد تحليل الأشعة السينية بنية عديد الببتيد. البروتين التأكيد الرئيسي لنظرية عديد الببتيد للبروتين هو إمكانية تصنيع عديدات الببتيد والبروتينات من البروتينات المعروفة بالفعل. البروتين (الأنسولين، الليزوزيم، النوى الريبية) بالطرق الكيميائية.
الدليل التجريبي للنظرية l l l تحتوي البروتينات الطبيعية على عدد صغير من المجموعات المعايرة – COOH و –NH 2. أثناء عملية التحلل المائي للبروتين، يتم إنشاء عدد متكافئ من المجموعات المعايرة – COOH و –NH 2 تحت تأثير البروتينات. في وجود إنزيمات معينة، يتم تقسيم البروتينات إلى أجزاء محددة بدقة (عديدات الببتيد). ويؤكد تحليل الأشعة السينية بنية عديد الببتيد. البروتين التأكيد الرئيسي لنظرية عديد الببتيد للبروتين هو إمكانية تصنيع عديدات الببتيد والبروتينات من البروتينات المعروفة بالفعل. البروتين (الأنسولين، الليزوزيم، النوى الريبية) بالطرق الكيميائية.

 السمات الهيكلية لرمح الببتيد l l رابط الببتيد أقصر بنسبة 10٪ تقريبًا من رابط –C – N– وله طابع الارتباط “المغمور بشكل متكرر” –C = N– بواسطة L. Pauling و R. Corey ، والذي كان تم تطويره في 1948-1955 ص. اشرح الطبيعة الخاصة للرابطة C-N عن طريق "الرنين" بين الشكلين
السمات الهيكلية لرمح الببتيد l l رابط الببتيد أقصر بنسبة 10٪ تقريبًا من رابط –C – N– وله طابع الارتباط “المغمور بشكل متكرر” –C = N– بواسطة L. Pauling و R. Corey ، والذي كان تم تطويره في 1948-1955 ص. اشرح الطبيعة الخاصة للرابطة C-N عن طريق "الرنين" بين الشكلين
 السمات الهيكلية لرمح الببتيد l l l وبعبارة أخرى، في البروتينات والببتيدات، غالبًا ما تكون روابط C-N متعددة من خلال تفاعل زوج الإلكترونات الوحيد لذرة النيتروجين مع النظام الإلكتروني لمجموعة الكربونيل، مما يؤدي إلى صعوبة تكوين tannya navkolo zvjazku C–N Zazvichay peptide zv' يكون تكوين trans أكثر وضوحًا من تكوين cis بمقدار 2.6 كيلو كالوري/مول (10.878 كيلوجول/مول)، نظرًا لأن قرب ذرات الكربون في تكوين cis يجعل من الصعب
السمات الهيكلية لرمح الببتيد l l l وبعبارة أخرى، في البروتينات والببتيدات، غالبًا ما تكون روابط C-N متعددة من خلال تفاعل زوج الإلكترونات الوحيد لذرة النيتروجين مع النظام الإلكتروني لمجموعة الكربونيل، مما يؤدي إلى صعوبة تكوين tannya navkolo zvjazku C–N Zazvichay peptide zv' يكون تكوين trans أكثر وضوحًا من تكوين cis بمقدار 2.6 كيلو كالوري/مول (10.878 كيلوجول/مول)، نظرًا لأن قرب ذرات الكربون في تكوين cis يجعل من الصعب
 السمات الهيكلية لرمح الببتيد l l l الدورانات ممكنة حول رابطتين بسيطتين (N–C وC–C 1)، المتجاورتين للذرة غير المتماثلة = 180 - شكل تحويلي)
السمات الهيكلية لرمح الببتيد l l l الدورانات ممكنة حول رابطتين بسيطتين (N–C وC–C 1)، المتجاورتين للذرة غير المتماثلة = 180 - شكل تحويلي)
 السمات الهيكلية لمشرط الببتيد l l نظرًا للتفاعل بين المدافعين في مشرط الببتيد، لا يمكنهم أن يأخذوا أي معنى - يُسمح لهم فقط بمناطق معينة منفصلة، مما يشير إلى النشاط النشط. بعض المطابقات للببتيد لانجوج كوتي لها علاقات متبادلة، فتغيير أحدهما أصعب من تغيير الآخر
السمات الهيكلية لمشرط الببتيد l l نظرًا للتفاعل بين المدافعين في مشرط الببتيد، لا يمكنهم أن يأخذوا أي معنى - يُسمح لهم فقط بمناطق معينة منفصلة، مما يشير إلى النشاط النشط. بعض المطابقات للببتيد لانجوج كوتي لها علاقات متبادلة، فتغيير أحدهما أصعب من تغيير الآخر

 L K. W. Linderstöm-Lang ميز 4 مستويات من تنظيم جزيئات البروتين - الهياكل الأولية والثانوية والثالثية والرباعية. على الرغم من أن هذه الفئات أصبحت قديمة، إلا أنها ستستمر في استخدامها في الوقت الحاضر.
L K. W. Linderstöm-Lang ميز 4 مستويات من تنظيم جزيئات البروتين - الهياكل الأولية والثانوية والثالثية والرباعية. على الرغم من أن هذه الفئات أصبحت قديمة، إلا أنها ستستمر في استخدامها في الوقت الحاضر.

 يتم تشفير البنية الأولية للبروتين l تسلسل بقايا الأحماض الأمينية في انسيت متعدد الببتيد بواسطة الجين الهيكلي لهذا البروتين ويحتوي على كل ما هو ضروري للتنظيم الذاتي لبنيته الواسعة. جميع البروتينات تتنافس على خاصيتها مع هذه البنية الأولية، ولا يقتصر العدد المحتمل لهذه الهياكل على عدد أنواع البروتينات المختلفة في جميع أنواع الكائنات الحية.حوالي 1010-1012
يتم تشفير البنية الأولية للبروتين l تسلسل بقايا الأحماض الأمينية في انسيت متعدد الببتيد بواسطة الجين الهيكلي لهذا البروتين ويحتوي على كل ما هو ضروري للتنظيم الذاتي لبنيته الواسعة. جميع البروتينات تتنافس على خاصيتها مع هذه البنية الأولية، ولا يقتصر العدد المحتمل لهذه الهياكل على عدد أنواع البروتينات المختلفة في جميع أنواع الكائنات الحية.حوالي 1010-1012
 من المستحيل فهم الوظيفة البيولوجية والآلية الجزيئية للنشاط الفسيولوجي للبروتين دون معرفة تفصيلية بوجوده. إن دراسة البنية الأولية للبروتينات "المطفرة" تجعل من الممكن فهم طبيعة الأمراض الركودية على المستوى الجزيئي.
من المستحيل فهم الوظيفة البيولوجية والآلية الجزيئية للنشاط الفسيولوجي للبروتين دون معرفة تفصيلية بوجوده. إن دراسة البنية الأولية للبروتينات "المطفرة" تجعل من الممكن فهم طبيعة الأمراض الركودية على المستوى الجزيئي.
 طرق تحديد البنية الأولية يهدف تحضير البروتين قبل تحليل البنية الأولية إلى التقليل إلى أدنى حد من ضخ مستويات أخرى أعلى من تنظيمه، وبعبارة أخرى، قد يكون موضوع التحليل مضطربًا في هذا البروتين دون أي روابط تساهمية عرضية (على سبيل المثال، ثاني كبريتيد) بحيث تكون جميع الروابط الببتيدية متاحة لكل من الكواشف الكيميائية والإنزيمات
طرق تحديد البنية الأولية يهدف تحضير البروتين قبل تحليل البنية الأولية إلى التقليل إلى أدنى حد من ضخ مستويات أخرى أعلى من تنظيمه، وبعبارة أخرى، قد يكون موضوع التحليل مضطربًا في هذا البروتين دون أي روابط تساهمية عرضية (على سبيل المثال، ثاني كبريتيد) بحيث تكون جميع الروابط الببتيدية متاحة لكل من الكواشف الكيميائية والإنزيمات
 البروتين هو المسؤول أولاً عن كل عمليات تمسخ الطبيعة العميقة وفقدان البنية الرباعية والثالثية والثانوية إن أمكن.
البروتين هو المسؤول أولاً عن كل عمليات تمسخ الطبيعة العميقة وفقدان البنية الرباعية والثالثية والثانوية إن أمكن.
 لمنع تكوين روابط ثاني كبريتيد، امنع مجموعات السلفهيدريل مرة أخرى باستخدام حمض اليودوكتيك الزائد: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI
لمنع تكوين روابط ثاني كبريتيد، امنع مجموعات السلفهيدريل مرة أخرى باستخدام حمض اليودوكتيك الزائد: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI
 بعد ذلك، يتم إجراء التحلل المائي الانتقائي للبروتين المشوه لإزالة نظام الببتيد الذي يتداخل. يتم إجراء التحلل المائي باستخدام الإنزيمات: l l l التربسين (الارتباطات التي تم إنشاؤها بواسطة مجموعات الكربوكسيل من الأحماض الأمينية العطرية) والكيموتربسين (الارتباطات التي تم إنشاؤها بواسطة مجموعات -COOH من الأحماض الأمينية العطرية) وغيرها ؛ الكواشف الكيميائية: البروموسيان Br - C N (المواد الرابطة المركبة بمجموعات - COOH met) وغيرها.
بعد ذلك، يتم إجراء التحلل المائي الانتقائي للبروتين المشوه لإزالة نظام الببتيد الذي يتداخل. يتم إجراء التحلل المائي باستخدام الإنزيمات: l l l التربسين (الارتباطات التي تم إنشاؤها بواسطة مجموعات الكربوكسيل من الأحماض الأمينية العطرية) والكيموتربسين (الارتباطات التي تم إنشاؤها بواسطة مجموعات -COOH من الأحماض الأمينية العطرية) وغيرها ؛ الكواشف الكيميائية: البروموسيان Br - C N (المواد الرابطة المركبة بمجموعات - COOH met) وغيرها.
 l l l المرحلة الأولى - تجزئة الببتيدات المعزولة - يتم تنفيذها بطرق كروماتوغرافيا، المرحلة المتبقية - تحديد البنية الأولية l تبدأ بأهمية الوزن الجزيئي، تكوين الأحماض الأمينية، N- والأحماض الأمينية الطرفية. عامل آخر واختيار نوع آخر مختلف من المجموعة الأولى من شظايا الببتيد، والتي يتم تقسيمها وتحليلها لتحديد ترتيب توزيع الأحماض الأمينية في البروتين
l l l المرحلة الأولى - تجزئة الببتيدات المعزولة - يتم تنفيذها بطرق كروماتوغرافيا، المرحلة المتبقية - تحديد البنية الأولية l تبدأ بأهمية الوزن الجزيئي، تكوين الأحماض الأمينية، N- والأحماض الأمينية الطرفية. عامل آخر واختيار نوع آخر مختلف من المجموعة الأولى من شظايا الببتيد، والتي يتم تقسيمها وتحليلها لتحديد ترتيب توزيع الأحماض الأمينية في البروتين

 واو طريقة سانجر ل ش 1945 ص. طور عالم الكيمياء الحيوية الإنجليزي F. Sanger إحدى الطرق الأولى لتحديد الأحماض الأمينية الطرفية N
واو طريقة سانجر ل ش 1945 ص. طور عالم الكيمياء الحيوية الإنجليزي F. Sanger إحدى الطرق الأولى لتحديد الأحماض الأمينية الطرفية N
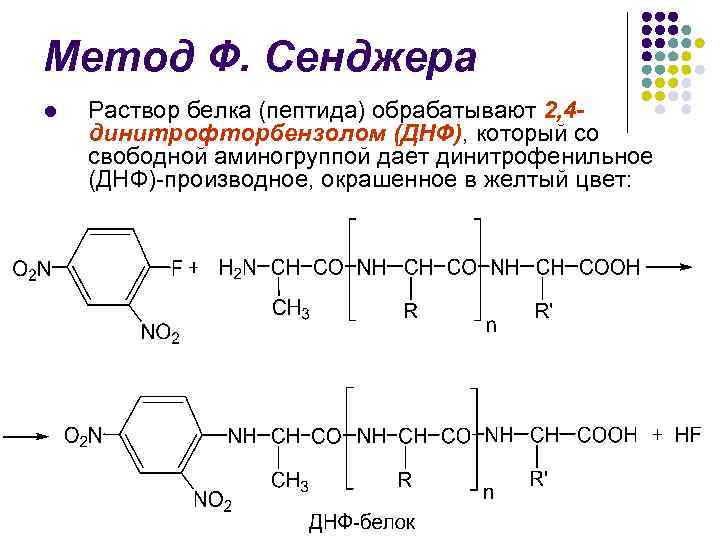 F. طريقة سانجر ل يتم معالجة البروتين (الببتيد) بـ 2،4 دينيتروفلوروبنزين (DNF) والذي يعطي من المجموعة الأمينية الحرة دينيتروفينيل (DNF) المحتوي على اللون الأصفر:
F. طريقة سانجر ل يتم معالجة البروتين (الببتيد) بـ 2،4 دينيتروفلوروبنزين (DNF) والذي يعطي من المجموعة الأمينية الحرة دينيتروفينيل (DNF) المحتوي على اللون الأصفر:
 F. طريقة سانجر l l يؤدي التحلل المائي للحمض الهجومي (5.7 N HCl) إلى تدمير الروابط الببتيدية وإنشاء حمض أميني N- مثل DNP.
F. طريقة سانجر l l يؤدي التحلل المائي للحمض الهجومي (5.7 N HCl) إلى تدمير الروابط الببتيدية وإنشاء حمض أميني N- مثل DNP.
 طريقة دانسيلني في روزروبليني عام 1963. علماء الكيمياء الحيوية الإنجليز دبليو جراي وبي هارتلي
طريقة دانسيلني في روزروبليني عام 1963. علماء الكيمياء الحيوية الإنجليز دبليو جراي وبي هارتلي
 طريقة دانسيل l المرحلة الأولى - تفاعل كلوريد الدانسيل (1-ثنائي ميثيل أمينونافثالين-5-سلفوكلوريد) مع ببتيد مجموعة أمينية غير بروتونية (أو بروتين) مع ببتيد دانسيل معدل (ببتيد DNS)
طريقة دانسيل l المرحلة الأولى - تفاعل كلوريد الدانسيل (1-ثنائي ميثيل أمينونافثالين-5-سلفوكلوريد) مع ببتيد مجموعة أمينية غير بروتونية (أو بروتين) مع ببتيد دانسيل معدل (ببتيد DNS)

 أقوى طريقة لإزالة بروتينات DNS تخضع للتحلل المائي في وجود 5،7 ن. حمض الهيدروكلوريك، عند 105 على مدى 12-16 سنة، وبعد ذلك تم فصل الحمض الأميني DNS، وتم التعرف عليه عن طريق التألق في تبادلات الأشعة فوق البنفسجية
أقوى طريقة لإزالة بروتينات DNS تخضع للتحلل المائي في وجود 5،7 ن. حمض الهيدروكلوريك، عند 105 على مدى 12-16 سنة، وبعد ذلك تم فصل الحمض الأميني DNS، وتم التعرف عليه عن طريق التألق في تبادلات الأشعة فوق البنفسجية


 طريقة S. Akabori l l عندما يتم تسخين الببتيد (أو البروتين) مع الهيدرازين اللامائي عند 100-120 درجة مئوية، يتم تحلل روابط الببتيد مع الأحماض الأمينية للهيدرازيدات، ويتم فقدان الحمض الأميني الطرفي C في المظهر. يمكن رؤيتها من الخليط وتحديد:
طريقة S. Akabori l l عندما يتم تسخين الببتيد (أو البروتين) مع الهيدرازين اللامائي عند 100-120 درجة مئوية، يتم تحلل روابط الببتيد مع الأحماض الأمينية للهيدرازيدات، ويتم فقدان الحمض الأميني الطرفي C في المظهر. يمكن رؤيتها من الخليط وتحديد:

 طريقة P. Edman l تم تحليل طريقة تحلل البولي ببتيد بمساعدة فينيل إيزوثيوسيونات (FITC) من قبل الكيميائي السويدي P. V. Edman في 1950-1956. l تسمح طريقة Edman بالإزالة المتسقة لبقايا الأحماض الأمينية N-terminal من شكل فينيل ثيوهيدانتوين (PTH) l تتضمن دورة تحلل الجلد 3 مراحل
طريقة P. Edman l تم تحليل طريقة تحلل البولي ببتيد بمساعدة فينيل إيزوثيوسيونات (FITC) من قبل الكيميائي السويدي P. V. Edman في 1950-1956. l تسمح طريقة Edman بالإزالة المتسقة لبقايا الأحماض الأمينية N-terminal من شكل فينيل ثيوهيدانتوين (PTH) l تتضمن دورة تحلل الجلد 3 مراحل
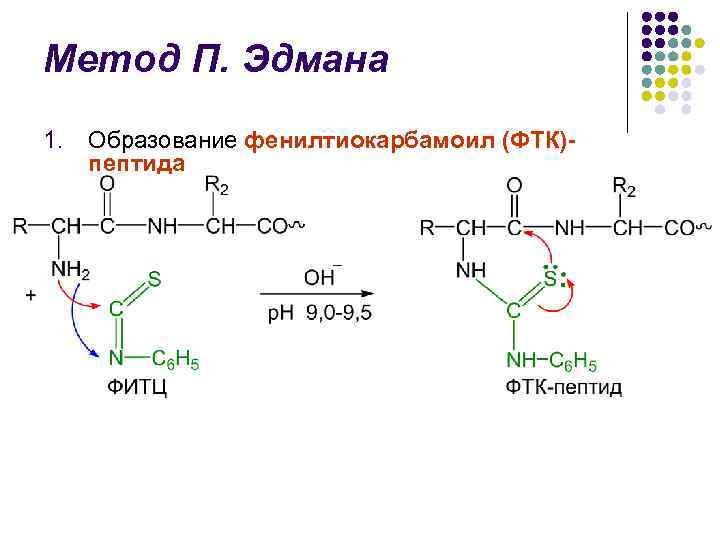



 طريقة إدمان ل ل ل تحديد فواصل PTG يتم في البداية بطريقة إدمان، خلال الثلاث ساعات الماضية تم إجراء كروماتوغرافيا على الورق، ثم كروماتوغرافيا الكرة الدقيقة على هلام السيليكا والبولي أميد، كروماتوغرافيا الغاز الطبيعي الأصلي إنجازات كبيرة في مجال التركيب الهيكلي بحث. جهاز تسلسل Edman و J. Begg (من التسلسل الإنجليزي - التسلسل) - جهاز يقوم تلقائيًا بكفاءة عالية بفصل بقايا الأحماض الأمينية N-terminal باستخدام طريقة Edman
طريقة إدمان ل ل ل تحديد فواصل PTG يتم في البداية بطريقة إدمان، خلال الثلاث ساعات الماضية تم إجراء كروماتوغرافيا على الورق، ثم كروماتوغرافيا الكرة الدقيقة على هلام السيليكا والبولي أميد، كروماتوغرافيا الغاز الطبيعي الأصلي إنجازات كبيرة في مجال التركيب الهيكلي بحث. جهاز تسلسل Edman و J. Begg (من التسلسل الإنجليزي - التسلسل) - جهاز يقوم تلقائيًا بكفاءة عالية بفصل بقايا الأحماض الأمينية N-terminal باستخدام طريقة Edman
 لتحديد بنية الببتيدات والبروتينات ، يمكن تكثيف ما يلي: l l الإنزيمات التي تحفز التخلص من بقايا الأحماض الأمينية الطرفية N و C - الكربوكسيببتيداز الأميني ، الطرق الفيزيائية والكيميائية ، التحليل الطيفي ، قياس الطيف الكتلي يو
لتحديد بنية الببتيدات والبروتينات ، يمكن تكثيف ما يلي: l l الإنزيمات التي تحفز التخلص من بقايا الأحماض الأمينية الطرفية N و C - الكربوكسيببتيداز الأميني ، الطرق الفيزيائية والكيميائية ، التحليل الطيفي ، قياس الطيف الكتلي يو
 إن تحليل البيانات المتعلقة بالبنية الأولية يجعل من الممكن تطوير المزيد من المبادئ الأساسية، ويتم ضمان استقرار البنية الأولية بشكل رئيسي عن طريق الروابط الببتيدية؛ احتمالية المشاركة وعدد قليل من روابط ثاني كبريتيد في رمح البولي ببتيد، قد يتم الكشف عن مجموعات مختلفة من الأحماض الأمينية.في البولي ببتيد، يتم الكشف عن جميع ثنائيات الببتيد المحتملة.
إن تحليل البيانات المتعلقة بالبنية الأولية يجعل من الممكن تطوير المزيد من المبادئ الأساسية، ويتم ضمان استقرار البنية الأولية بشكل رئيسي عن طريق الروابط الببتيدية؛ احتمالية المشاركة وعدد قليل من روابط ثاني كبريتيد في رمح البولي ببتيد، قد يتم الكشف عن مجموعات مختلفة من الأحماض الأمينية.في البولي ببتيد، يتم الكشف عن جميع ثنائيات الببتيد المحتملة.
 يتيح لنا تحليل البيانات المتعلقة بالبنية الأولية تطوير المبادئ الأساسية التالية: l l يتميز جلد البروتين المتجانس الفردي ببنية أولية فريدة من نوعها؛ في كثير من الأحيان، استبدال الأحماض الأمينية لا يؤدي فقط إلى تغييرات هيكلية، ولكن أيضا إلى تغييرات في الخصائص الفيزيائية والكيميائية والوظائف البيولوجية. يعد مبدأ التشابه الهيكلي هذا أكثر شيوعًا بالنسبة للإنزيمات منخفضة التحلل للبروتين - التربسين والكيموتربسين وما إلى ذلك.
يتيح لنا تحليل البيانات المتعلقة بالبنية الأولية تطوير المبادئ الأساسية التالية: l l يتميز جلد البروتين المتجانس الفردي ببنية أولية فريدة من نوعها؛ في كثير من الأحيان، استبدال الأحماض الأمينية لا يؤدي فقط إلى تغييرات هيكلية، ولكن أيضا إلى تغييرات في الخصائص الفيزيائية والكيميائية والوظائف البيولوجية. يعد مبدأ التشابه الهيكلي هذا أكثر شيوعًا بالنسبة للإنزيمات منخفضة التحلل للبروتين - التربسين والكيموتربسين وما إلى ذلك.

 في الاندماج غير القطبي، تصبح طاقة الرباط المائي –CO НN– قريبة من 16.7 كيلوجول/مول، ويؤدي التحول في قطبية التيار الأوسط إلى تقليل هذه الطاقة
في الاندماج غير القطبي، تصبح طاقة الرباط المائي –CO НN– قريبة من 16.7 كيلوجول/مول، ويؤدي التحول في قطبية التيار الأوسط إلى تقليل هذه الطاقة
 التفاعلات المتبادلة الكارهة للماء لها طبيعة انتروبية بسبب دخول المدافعين غير القطبيين إلى الماء ومحاولة الحد من اتصالهم به، على سبيل المثال، الماء لا يجدد بنيته الهيكلية وكما لو كانت مجموعة بريموس هناك شفعاء فيها الكتلة التي تحتوي على الحد الأدنى من الطاقة. فائض
التفاعلات المتبادلة الكارهة للماء لها طبيعة انتروبية بسبب دخول المدافعين غير القطبيين إلى الماء ومحاولة الحد من اتصالهم به، على سبيل المثال، الماء لا يجدد بنيته الهيكلية وكما لو كانت مجموعة بريموس هناك شفعاء فيها الكتلة التي تحتوي على الحد الأدنى من الطاقة. فائض

 تتكون تفاعلات فان دير فالس من قوى التشتت وجاذبية الذرات وقوى التكيف المتبادل لأغلفتها الإلكترونية، وتكون مساهمة الطاقة في ملامسة الجلد صغيرة (
تتكون تفاعلات فان دير فالس من قوى التشتت وجاذبية الذرات وقوى التكيف المتبادل لأغلفتها الإلكترونية، وتكون مساهمة الطاقة في ملامسة الجلد صغيرة (
 التفاعلات الأيونية أو الكهروستاتيكية تفاعلات المجموعات المشحونة l تفاعلات المجموعات الأيونية التي تولدها الروابط الملحية يمكن أن تصل طاقة الروابط الملحية إلى سطح كاره للماء 41.9 كيلو جول/مول، لكن كميتها صغيرة جدًا. تفاعلات ثنائية أيونية وثنائية القطب مماثلة
التفاعلات الأيونية أو الكهروستاتيكية تفاعلات المجموعات المشحونة l تفاعلات المجموعات الأيونية التي تولدها الروابط الملحية يمكن أن تصل طاقة الروابط الملحية إلى سطح كاره للماء 41.9 كيلو جول/مول، لكن كميتها صغيرة جدًا. تفاعلات ثنائية أيونية وثنائية القطب مماثلة
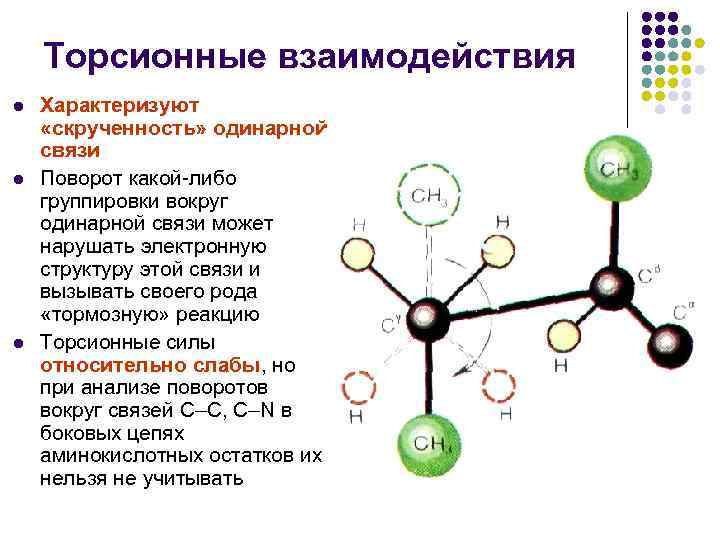 تفاعلات الالتواء l l l تميز "التواء" رابطة واحدة. إن دوران أي تجمع حول رابطة واحدة يمكن أن يدمر البنية الإلكترونية للرابطة الخاصة بها ويؤدي إلى نوع من رد فعل "الهالم". من الواضح أن تفاعل الالتواء والقوى ضعيفة، ولكن عندما لا يمكن تجنب تحليل المنعطفات حول الأربطة С–С، С–N من نفايات الأحماض الأمينية الزائدة.
تفاعلات الالتواء l l l تميز "التواء" رابطة واحدة. إن دوران أي تجمع حول رابطة واحدة يمكن أن يدمر البنية الإلكترونية للرابطة الخاصة بها ويؤدي إلى نوع من رد فعل "الهالم". من الواضح أن تفاعل الالتواء والقوى ضعيفة، ولكن عندما لا يمكن تجنب تحليل المنعطفات حول الأربطة С–С، С–N من نفايات الأحماض الأمينية الزائدة.

 البنية الثانوية للبروتين... سوف تقوم بتوسيع واسع النطاق للأجزاء المستديرة من المشرط متعدد الببتيد بدون نوع وشكل جذور الأحماض الأمينية. يتم إنشاؤه عن طريق تفاعل الروابط المائية بين مجموعات الببتيد من كل من المشرط الواحد والمجموعات المختلفة. لانتسيوغيف
البنية الثانوية للبروتين... سوف تقوم بتوسيع واسع النطاق للأجزاء المستديرة من المشرط متعدد الببتيد بدون نوع وشكل جذور الأحماض الأمينية. يتم إنشاؤه عن طريق تفاعل الروابط المائية بين مجموعات الببتيد من كل من المشرط الواحد والمجموعات المختلفة. لانتسيوغيف
 البنية الثانوية للبروتين كل قسم من جزيء البروتين له بنية ثانوية، ويرى البعض البنية الثانوية للعناصر الأكثر دورية: - الحلزون و - البنية. ومع ذلك، في البروتينات، تصبح الأقسام، التي يتم تنظيمها، أكثر دقة في بطريقة لطيفة، على الرغم من أن هيكلها الواسع لا ينتقم من أحد من وقت لآخر، فهو عبارة متكررة ومنتظمة. قبلهم، نحن نفهم تمامًا مفهوم البنية الثانوية، هناك نوعان من الهياكل الثانوية: منتظمة وغير منتظمة، ولا ينطبق مفهوم البنية الثانوية على جميع جزيئات البروتين ككل، ولكن على بضعة أقسام أكبر من الأرضية. لانسيت ايبيبتيد
البنية الثانوية للبروتين كل قسم من جزيء البروتين له بنية ثانوية، ويرى البعض البنية الثانوية للعناصر الأكثر دورية: - الحلزون و - البنية. ومع ذلك، في البروتينات، تصبح الأقسام، التي يتم تنظيمها، أكثر دقة في بطريقة لطيفة، على الرغم من أن هيكلها الواسع لا ينتقم من أحد من وقت لآخر، فهو عبارة متكررة ومنتظمة. قبلهم، نحن نفهم تمامًا مفهوم البنية الثانوية، هناك نوعان من الهياكل الثانوية: منتظمة وغير منتظمة، ولا ينطبق مفهوم البنية الثانوية على جميع جزيئات البروتين ككل، ولكن على بضعة أقسام أكبر من الأرضية. لانسيت ايبيبتيد
 البنية الثانوية للبروتين l يتم الإشارة إلى التفاعلات التي تلعب الدور الأكثر أهمية في البنية الثانوية المتكونة من خلال: l l خصائص الرابطة الببتيدية والتفاعلات الاستاتيكية (- و-القطع) الدور الرئيسي تلعبه الروابط المائية بين البروتينات. مجموعات الببتيد والتي تتكرر بشكل دوري في لانكوس
البنية الثانوية للبروتين l يتم الإشارة إلى التفاعلات التي تلعب الدور الأكثر أهمية في البنية الثانوية المتكونة من خلال: l l خصائص الرابطة الببتيدية والتفاعلات الاستاتيكية (- و-القطع) الدور الرئيسي تلعبه الروابط المائية بين البروتينات. مجموعات الببتيد والتي تتكرر بشكل دوري في لانكوس
 تثبيت الهيكل الثانوي لتشكيل روابط الماء ل حقن البروتين الزائد بالماء يمكن أن يتنافس الماء بشكل فعال لتشكيل روابط الماء: يتم تقليل ضخ الماء عند تشكيل بنية بروتينية واسعة مدمجة ونمو وبدلاً من روابط الببتيد، زيادة توافق تفاعلاتها وبالتالي استقرار البنية الثانوية ليقع في البنية الثلاثية المدمجة
تثبيت الهيكل الثانوي لتشكيل روابط الماء ل حقن البروتين الزائد بالماء يمكن أن يتنافس الماء بشكل فعال لتشكيل روابط الماء: يتم تقليل ضخ الماء عند تشكيل بنية بروتينية واسعة مدمجة ونمو وبدلاً من روابط الببتيد، زيادة توافق تفاعلاتها وبالتالي استقرار البنية الثانوية ليقع في البنية الثلاثية المدمجة

 -دوامة ل في صخور الخمسينيات من القرن العشرين. L. Pauling و R. Corey، استنادًا إلى بيانات حول بنية بلورات الأحماض الأمينية والببتيدات البسيطة، نظروا في المطابقات الدورية المحتملة للبولي ببتيد لانجوج وتوصلوا إلى استنتاج مفاده أن الهيكل الأكثر تجانسًا تم تسميته بـ -spi رالي
-دوامة ل في صخور الخمسينيات من القرن العشرين. L. Pauling و R. Corey، استنادًا إلى بيانات حول بنية بلورات الأحماض الأمينية والببتيدات البسيطة، نظروا في المطابقات الدورية المحتملة للبولي ببتيد لانجوج وتوصلوا إلى استنتاج مفاده أن الهيكل الأكثر تجانسًا تم تسميته بـ -spi رالي
 - حلزوني يعتمد اختيار الموضع على المعايير التالية: 1. إنشاء بنية مدمجة محكمة الغلق بدون ذرات فارغة ومبالغ فيها 2. الحد الأقصى لتشبع الهيكل بالروابط المائية من هذه المادة الكيميائية، بحيث كانت هندستها قريبة من الخطية 3 Dotrimannya بين الناهضات الذرية والزوايا وتسامح الببتيدات l
- حلزوني يعتمد اختيار الموضع على المعايير التالية: 1. إنشاء بنية مدمجة محكمة الغلق بدون ذرات فارغة ومبالغ فيها 2. الحد الأقصى لتشبع الهيكل بالروابط المائية من هذه المادة الكيميائية، بحيث كانت هندستها قريبة من الخطية 3 Dotrimannya بين الناهضات الذرية والزوايا وتسامح الببتيدات l
 - الحلزون l في أذهان هؤلاء الأشخاص، يمكن أن يكون هناك دوامة يمينية ويسارية، ولكن بالنسبة لليمين، تظهر اللولبية بشكل أكثر بروزًا من اليسار، نظرًا لأن رمح الببتيد يتكون من الأحماض الأمينية L
- الحلزون l في أذهان هؤلاء الأشخاص، يمكن أن يكون هناك دوامة يمينية ويسارية، ولكن بالنسبة لليمين، تظهر اللولبية بشكل أكثر بروزًا من اليسار، نظرًا لأن رمح الببتيد يتكون من الأحماض الأمينية L
 -Spiral l l يتطور الهيكل الحلزوني عندما تدور جميع جوانب المشرط متعدد الببتيد حول الأربطة البسيطة (i) التي لها نفس القيمة والعلامة (قريبة من 60)، مما يؤدي إلى التواء المشرط خطوة بخطوة عند هذه النقطة الجذور عبارة عن الكثير من الأحماض غير المشبعة التي تظهر على محيط دوامة الأسطوانة المذابة ويمكن، اعتمادًا على طبيعتها، ضمان الطبيعة الكارهة للماء أو المحبة للماء للسطح الأسطواني
-Spiral l l يتطور الهيكل الحلزوني عندما تدور جميع جوانب المشرط متعدد الببتيد حول الأربطة البسيطة (i) التي لها نفس القيمة والعلامة (قريبة من 60)، مما يؤدي إلى التواء المشرط خطوة بخطوة عند هذه النقطة الجذور عبارة عن الكثير من الأحماض غير المشبعة التي تظهر على محيط دوامة الأسطوانة المذابة ويمكن، اعتمادًا على طبيعتها، ضمان الطبيعة الكارهة للماء أو المحبة للماء للسطح الأسطواني
 - المعلمات الهندسية للدوامة: l l l l radius r = 2.3 Å (0.23 nm) ارتفاع الحلزوني (الإزاحة) لكل 1 زائد d = 1.5 Å حافة اللولب (فترة الهوية) P = 5.4 Å 1 دورة للدوامة، الشكل 3، 6 زيادات من الأحماض الأمينية، جميعها – روابط C=O مباشرة إلى الأمام، إلى النهاية C، والمجموعات –N–H في الخلف بشكل مستقيم.في الحلزون، ترتبط مجموعة الجلد –NH بواسطة وصلة مائية إلى مجموعة –CO من الحمض الأميني الرابع أكثر من اللازم. زنيوغو (مكالمة 5 1)
- المعلمات الهندسية للدوامة: l l l l radius r = 2.3 Å (0.23 nm) ارتفاع الحلزوني (الإزاحة) لكل 1 زائد d = 1.5 Å حافة اللولب (فترة الهوية) P = 5.4 Å 1 دورة للدوامة، الشكل 3، 6 زيادات من الأحماض الأمينية، جميعها – روابط C=O مباشرة إلى الأمام، إلى النهاية C، والمجموعات –N–H في الخلف بشكل مستقيم.في الحلزون، ترتبط مجموعة الجلد –NH بواسطة وصلة مائية إلى مجموعة –CO من الحمض الأميني الرابع أكثر من اللازم. زنيوغو (مكالمة 5 1)
 عدد المقاطع الحلزونية في البروتينات الكروية صغير بشكل ملحوظ (5 - 15 فائض من الأحماض الأمينية، 3 - 4 دورات من الحلزون)، في البروتينات الليفية - ممتدة بشكل غني. يرجى الاتصال بالأماكن التي يتم فيها تشغيل الفائض بحيث يمكن مقاطعة الأنظمة. عند هذه النقطة تنحسر الدوامة بأكملها لمدة 20-30
عدد المقاطع الحلزونية في البروتينات الكروية صغير بشكل ملحوظ (5 - 15 فائض من الأحماض الأمينية، 3 - 4 دورات من الحلزون)، في البروتينات الليفية - ممتدة بشكل غني. يرجى الاتصال بالأماكن التي يتم فيها تشغيل الفائض بحيث يمكن مقاطعة الأنظمة. عند هذه النقطة تنحسر الدوامة بأكملها لمدة 20-30

 -الهيكل ل ل اقترحه دبليو تي أستبري في عام 1941. على أساس الدراسات الهيكلية بالأشعة السينية - الكيراتين. بعد 10 سنوات، أثبت L. Pauling وR. Corey أن الهيكل، أو "الصفيحة القابلة للطي"، يرجع إلى استقرار الأربطة المائية بين الزعانف المرتبطة بالنسيج المتعرج المنسوج. الأربطة الببتيدية
-الهيكل ل ل اقترحه دبليو تي أستبري في عام 1941. على أساس الدراسات الهيكلية بالأشعة السينية - الكيراتين. بعد 10 سنوات، أثبت L. Pauling وR. Corey أن الهيكل، أو "الصفيحة القابلة للطي"، يرجع إلى استقرار الأربطة المائية بين الزعانف المرتبطة بالنسيج المتعرج المنسوج. الأربطة الببتيدية




 - البنية l l عدد فائض الأحماض الأمينية في جزء من رمح الببتيد، الذي يشكل البنية، يتراوح من 3 إلى 8. البنية الممتدة، التي تسمى الكرة، أو الورقة المطوية، تتكون في أغلب الأحيان من 2- 6 وخزات، وأحيانا تصل إلى 10
- البنية l l عدد فائض الأحماض الأمينية في جزء من رمح الببتيد، الذي يشكل البنية، يتراوح من 3 إلى 8. البنية الممتدة، التي تسمى الكرة، أو الورقة المطوية، تتكون في أغلب الأحيان من 2- 6 وخزات، وأحيانا تصل إلى 10
 - البنية l l l تظهر المجموعات الحمضية الزائدة من الأحماض الأمينية، عند تكوين البنية، من خلال الجوانب المختلفة لسطحها، والسطح نفسه له شكل مطوي، ويتم تحديد الطيات بواسطة ذرات الكربون. تأتي منها، ويتم إنشاء مشط. يتيح لك هذا تشكيل وتمديد الأسطح الطويلة المملوءة بنفس النوع (على سبيل المثال، الكارهة للماء) من الجذور البيولوجية للسطح الكاره للماء للكرة المطوية، والتي تتفاعل مع بعضها البعض أو مع الحواف الكارهة للماء للحلزونات ، تشارك في النوى الجزيئية الداخلية الكارهة للماء، والتي من السهل تثبيت بنية البروتين
- البنية l l l تظهر المجموعات الحمضية الزائدة من الأحماض الأمينية، عند تكوين البنية، من خلال الجوانب المختلفة لسطحها، والسطح نفسه له شكل مطوي، ويتم تحديد الطيات بواسطة ذرات الكربون. تأتي منها، ويتم إنشاء مشط. يتيح لك هذا تشكيل وتمديد الأسطح الطويلة المملوءة بنفس النوع (على سبيل المثال، الكارهة للماء) من الجذور البيولوجية للسطح الكاره للماء للكرة المطوية، والتي تتفاعل مع بعضها البعض أو مع الحواف الكارهة للماء للحلزونات ، تشارك في النوى الجزيئية الداخلية الكارهة للماء، والتي من السهل تثبيت بنية البروتين

 -Vigin l l يتم تقديم كل من الحلزون والبنية في بروتينات كروية في مقاطع قصيرة، مما يعني أن جزءًا من البنية الثانوية للبروتين يقع على أنواع مختلفة من الحلقات، مما يسمح لك بتغيير اتجاه لانجوج الببتيد. عنصر هيكلي يسمح للبولي ببتيد بتدوير 180 درجة. 3 مجموعات من الببتيد - - فيجين مثبت بواسطة رابطة مائية واحدة تظهر دائمًا تقريبًا على سطح كرية البروتين، والتي غالبًا ما تلعب دورًا مهمًا في تفاعلاتها مع الجزيئات الأخرى
-Vigin l l يتم تقديم كل من الحلزون والبنية في بروتينات كروية في مقاطع قصيرة، مما يعني أن جزءًا من البنية الثانوية للبروتين يقع على أنواع مختلفة من الحلقات، مما يسمح لك بتغيير اتجاه لانجوج الببتيد. عنصر هيكلي يسمح للبولي ببتيد بتدوير 180 درجة. 3 مجموعات من الببتيد - - فيجين مثبت بواسطة رابطة مائية واحدة تظهر دائمًا تقريبًا على سطح كرية البروتين، والتي غالبًا ما تلعب دورًا مهمًا في تفاعلاتها مع الجزيئات الأخرى


 كشفت نتائج التحليل الهيكلي بالأشعة السينية أن طريقة طي جزيء البروتين يتم تحديدها من خلال تسلسل الأحماض الأمينية - gly، pro، asn Yakshcho مع 6 أحماض أمينية زائدة مجمعة 4 لدمج تكوين الحلزون، ثم هذه المجموعة هو مركز التصاعد. إذا اجتمعت 3 إضافات من 5 مجموعات لتكوين الهيكل، فإن هذا التجميع هو بذرة
كشفت نتائج التحليل الهيكلي بالأشعة السينية أن طريقة طي جزيء البروتين يتم تحديدها من خلال تسلسل الأحماض الأمينية - gly، pro، asn Yakshcho مع 6 أحماض أمينية زائدة مجمعة 4 لدمج تكوين الحلزون، ثم هذه المجموعة هو مركز التصاعد. إذا اجتمعت 3 إضافات من 5 مجموعات لتكوين الهيكل، فإن هذا التجميع هو بذرة

 البنية الثانوية (فوق الثانوية) للبروتينات l l يرتبط هذا المستوى من تنظيم جزيئات البروتين بوجود مجموعات من الهياكل الثانوية التي تتفاعل مع بعضها البعض.
البنية الثانوية (فوق الثانوية) للبروتينات l l يرتبط هذا المستوى من تنظيم جزيئات البروتين بوجود مجموعات من الهياكل الثانوية التي تتفاعل مع بعضها البعض.

 Superspiralized -spiralized l l يتم تشكيله عن طريق لف الملصق - اللوالب الحلزونية في البروتوفيبريل (-الكيراتين)، والتي تتحد في ميكروفيبريل - يتم ضغط اللوالب معًا بواسطة قوى فان دير فالس
Superspiralized -spiralized l l يتم تشكيله عن طريق لف الملصق - اللوالب الحلزونية في البروتوفيبريل (-الكيراتين)، والتي تتحد في ميكروفيبريل - يتم ضغط اللوالب معًا بواسطة قوى فان دير فالس

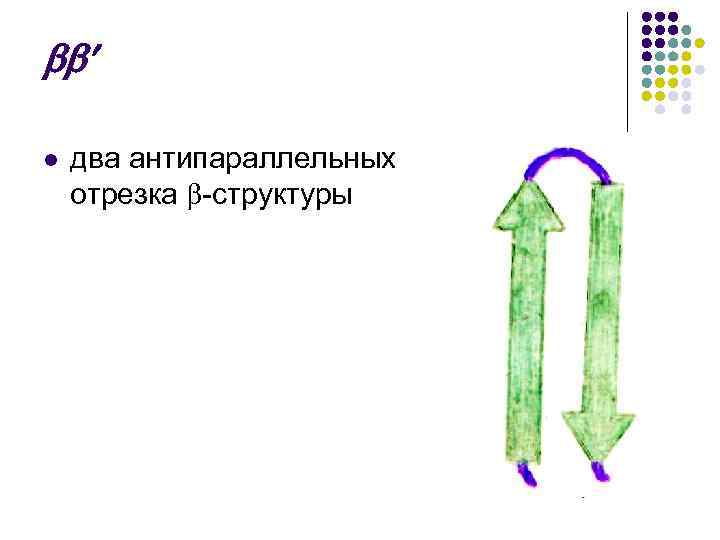
 x l حزام يتكون من كرتين متوازيتين بينهما مفاصل تشبه: l كرة غير منتظمة - مع
x l حزام يتكون من كرتين متوازيتين بينهما مفاصل تشبه: l كرة غير منتظمة - مع
 حزام x l، الذي يتكون من كرتين متوازيتين مع مفاصل بينهما، يبدو كما يلي: l l - اللوالب - مشرط وفقًا لروسمان. قطعتان متصلتان على التوالي تخلقان هيكل "الزخرفة اليونانية" -
حزام x l، الذي يتكون من كرتين متوازيتين مع مفاصل بينهما، يبدو كما يلي: l l - اللوالب - مشرط وفقًا لروسمان. قطعتان متصلتان على التوالي تخلقان هيكل "الزخرفة اليونانية" -
 حزام x l، الذي يتكون من كرتين متوازيتين بينهما مفاصل، يبدو كما يلي: l - هيكل - - - متعرج، "مفتاح الجوز"
حزام x l، الذي يتكون من كرتين متوازيتين بينهما مفاصل، يبدو كما يلي: l - هيكل - - - متعرج، "مفتاح الجوز"

 المجالات ومقاطع قصيرة واحدة تلو الأخرى من رمح متعدد الببتيد، والتي تسمى المقاطع المفصلية، يمكن تشكيل المجالات الوظيفية من واحد أو عدة مجالات هيكلية، وفي عدد من الإنزيمات، يتم فصل المركز النشط بين المجالات
المجالات ومقاطع قصيرة واحدة تلو الأخرى من رمح متعدد الببتيد، والتي تسمى المقاطع المفصلية، يمكن تشكيل المجالات الوظيفية من واحد أو عدة مجالات هيكلية، وفي عدد من الإنزيمات، يتم فصل المركز النشط بين المجالات



 3. + - البروتينات عبارة عن أقسام مكونة بشكل أساسي من الحلزونات، ومقاطع مكونة من كرات الأنسولين المضادة للتوازي
3. + - البروتينات عبارة عن أقسام مكونة بشكل أساسي من الحلزونات، ومقاطع مكونة من كرات الأنسولين المضادة للتوازي

 4. / - البروتينات l l - تتشكل اللوالب والهياكل على طول مسار Lancug. وتتمركز معظم الهياكل (المتوازية) في الجزء المركزي من الجزيء، حيث تنحني هذه الهياكل عند ظهور المروحة ("" "بنية ملتوية") ، مما يؤدي إلى إنشاء "قاعدة" صلبة ، والتي ترتبط بها أجزاء أخرى من الجزيء
4. / - البروتينات l l - تتشكل اللوالب والهياكل على طول مسار Lancug. وتتمركز معظم الهياكل (المتوازية) في الجزء المركزي من الجزيء، حيث تنحني هذه الهياكل عند ظهور المروحة ("" "بنية ملتوية") ، مما يؤدي إلى إنشاء "قاعدة" صلبة ، والتي ترتبط بها أجزاء أخرى من الجزيء


 المجالات l l l متوسط حجم المجال هو 100 – 150 إضافية، مما يشير إلى كريات يبلغ قطرها حوالي 2.5 نانومتر. هـ - تكوين البنية الثلاثية، والذي يحدث في المراحل النهائية من تكوين الكريات
المجالات l l l متوسط حجم المجال هو 100 – 150 إضافية، مما يشير إلى كريات يبلغ قطرها حوالي 2.5 نانومتر. هـ - تكوين البنية الثلاثية، والذي يحدث في المراحل النهائية من تكوين الكريات

 يُميِّز الهيكل الثلاثي للبروتين التوزيع الواسع للمقاطع المرتبة وغير المتبلورة في المشرط متعدد الببتيد ككل، والذي يرجع إلى تفاعل الجذور البيولوجية ويكمن في نوعها وشكلها.وهكذا، يصف الهيكل الثلاثي الترتيب المكاني لجميع جزيئات البروتين، حيث أنها مصنوعة من مادة واحدة متعددة الببتيد ترتبط بشكل مباشر بشكل جزيئات البروتين، والتي يمكن أن تختلف من خيطية إلى خيطية
يُميِّز الهيكل الثلاثي للبروتين التوزيع الواسع للمقاطع المرتبة وغير المتبلورة في المشرط متعدد الببتيد ككل، والذي يرجع إلى تفاعل الجذور البيولوجية ويكمن في نوعها وشكلها.وهكذا، يصف الهيكل الثلاثي الترتيب المكاني لجميع جزيئات البروتين، حيث أنها مصنوعة من مادة واحدة متعددة الببتيد ترتبط بشكل مباشر بشكل جزيئات البروتين، والتي يمكن أن تختلف من خيطية إلى خيطية
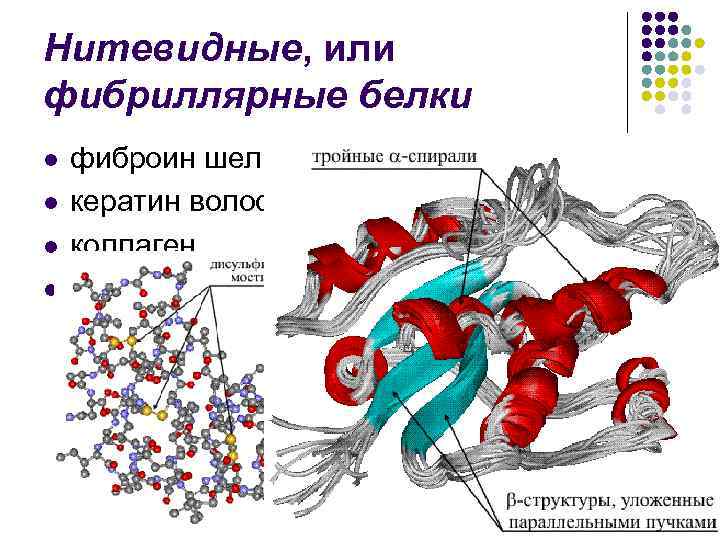 البروتينات الشبيهة بالخيوط أو الليفية لخياطة الكيراتين الليفي للشعر والقرون وتراكم الكولاجين وما إلى ذلك.
البروتينات الشبيهة بالخيوط أو الليفية لخياطة الكيراتين الليفي للشعر والقرون وتراكم الكولاجين وما إلى ذلك.

 البنية الثلاثية للبروتين هي أساس وظيفة البروتين، الأمر الذي يتطلب التنظيم المكاني الدقيق للمجموعات الكبيرة التي تم إنشاؤها في غياب تجاوزات الأحماض الأمينية. يجب إجراء عملية تمسخ البروتين حتى يفقد قدرته على العمل
البنية الثلاثية للبروتين هي أساس وظيفة البروتين، الأمر الذي يتطلب التنظيم المكاني الدقيق للمجموعات الكبيرة التي تم إنشاؤها في غياب تجاوزات الأحماض الأمينية. يجب إجراء عملية تمسخ البروتين حتى يفقد قدرته على العمل
 البنية الثلاثية للبروتين l l l يكمن استقرار البنية الثلاثية في نظام التفاعلات غير التساهمية في منتصف كرية البروتين (؟) يتم تثبيت بعض البروتينات بالإضافة إلى ذلك بواسطة روابط تساهمية - ثاني كبريتيد - Glanzug = RTln. K = Nchain – T S lanceug Nchain 0 (S
البنية الثلاثية للبروتين l l l يكمن استقرار البنية الثلاثية في نظام التفاعلات غير التساهمية في منتصف كرية البروتين (؟) يتم تثبيت بعض البروتينات بالإضافة إلى ذلك بواسطة روابط تساهمية - ثاني كبريتيد - Glanzug = RTln. K = Nchain – T S lanceug Nchain 0 (S
 البنية الثلاثية للبروتين l Spivvidnosti Chain i -T Schip تكمن في: l عدد الروابط غير التساهمية التي يتم تثبيتها عند ابتلاع جزيئات البروتين في الكرية، وبعد أن يعتمد عديد الببتيد على درجة الحرارة. Zi zrostannnyam T |T S| في وقت مبكر أو متأخر للتحرك | ن| يفقد هذا الهيكل المفتوح الاستقرار - تمسخ البروتين حرارياً
البنية الثلاثية للبروتين l Spivvidnosti Chain i -T Schip تكمن في: l عدد الروابط غير التساهمية التي يتم تثبيتها عند ابتلاع جزيئات البروتين في الكرية، وبعد أن يعتمد عديد الببتيد على درجة الحرارة. Zi zrostannnyam T |T S| في وقت مبكر أو متأخر للتحرك | ن| يفقد هذا الهيكل المفتوح الاستقرار - تمسخ البروتين حرارياً
 البنية الثلاثية للبروتين l l عندما يتم ابتلاع الكرية، تتغير إنتروبيا رمح الببتيد، ولكن في نفس الوقت هناك زيادة في إنتروبيا المنشئ - الماء، وهذا يلعب دورًا رئيسيًا في تثبيت البنية الثلاثية
البنية الثلاثية للبروتين l l عندما يتم ابتلاع الكرية، تتغير إنتروبيا رمح الببتيد، ولكن في نفس الوقت هناك زيادة في إنتروبيا المنشئ - الماء، وهذا يلعب دورًا رئيسيًا في تثبيت البنية الثلاثية
 البنية الثلاثية للبروتين l l الماء هو عامل تركيب غير مهم وعالي الترتيب (؟) يتم دمج المجموعات القطبية من البروتين بسهولة في الهياكل الشبيهة بالجليد في الماء، ويتم إدخال الروابط المائية منها. يشغلون مساحاتهم الفارغة التي أنشأتها الجزيئات. الماء، المرتبط بروابط مائية، كل هذا يوقف التدفق الفوضوي لجزيئات الماء، بحيث يكون هناك نظام، وتغير في إنتروبيا الماء
البنية الثلاثية للبروتين l l الماء هو عامل تركيب غير مهم وعالي الترتيب (؟) يتم دمج المجموعات القطبية من البروتين بسهولة في الهياكل الشبيهة بالجليد في الماء، ويتم إدخال الروابط المائية منها. يشغلون مساحاتهم الفارغة التي أنشأتها الجزيئات. الماء، المرتبط بروابط مائية، كل هذا يوقف التدفق الفوضوي لجزيئات الماء، بحيث يكون هناك نظام، وتغير في إنتروبيا الماء
 البنية الثلاثية للبروتين l l l إذا لم تتمكن أجزاء من المجموعات غير القطبية من البروتين من "الخروج بشكل كروي" من الجزيء، يتم تشكيل كرة، حيث يظهر جزء كبير (لا يقل عن ½) من المجموعات الكارهة للماء على اتصال مع ماء وهذا يستلزم تركيب جهات اتصال كارهة للماء، جهات اتصال فان دير فالس. القوى: هناك انخفاض في إنتروبيا البروتين. بعد تكوين الكرية، يحدث تدمير لجزء كبير من لزوجة الماء والتفاعلات الكارهة للماء لمجموعات البروتين مع جزيئات الماء. وتزداد فوضى الباقي، وتزداد فوضى الماء. يزداد انتروبيا الماء
البنية الثلاثية للبروتين l l l إذا لم تتمكن أجزاء من المجموعات غير القطبية من البروتين من "الخروج بشكل كروي" من الجزيء، يتم تشكيل كرة، حيث يظهر جزء كبير (لا يقل عن ½) من المجموعات الكارهة للماء على اتصال مع ماء وهذا يستلزم تركيب جهات اتصال كارهة للماء، جهات اتصال فان دير فالس. القوى: هناك انخفاض في إنتروبيا البروتين. بعد تكوين الكرية، يحدث تدمير لجزء كبير من لزوجة الماء والتفاعلات الكارهة للماء لمجموعات البروتين مع جزيئات الماء. وتزداد فوضى الباقي، وتزداد فوضى الماء. يزداد انتروبيا الماء

 البنية الرباعية للبروتين ... l l l تم العثور على البنية الرباعية في البروتينات قليلة القسيمات، التي تتكون جزيئاتها من اثنين أو أكثر من رماح عديد الببتيد المرتبطة بشكل غير تساهمي. مجموعة الكريات) تعمل كجزيء واحد، وفي نفس الوقت يحتفظ جلد الكريات المجمعة باستقلالية كبيرة.
البنية الرباعية للبروتين ... l l l تم العثور على البنية الرباعية في البروتينات قليلة القسيمات، التي تتكون جزيئاتها من اثنين أو أكثر من رماح عديد الببتيد المرتبطة بشكل غير تساهمي. مجموعة الكريات) تعمل كجزيء واحد، وفي نفس الوقت يحتفظ جلد الكريات المجمعة باستقلالية كبيرة.

 البنية الرباعية للبروتين l يمكن أن تكون الوحدات الفرعية التي تنشئ البنية الرباعية للبروتين مختلفة تمامًا عن وظائفها الفعلية - البروتينات غير المتجانسة، والتي تتيح لك الجمع بين عدد من الوظائف المترابطة في هيكل واحد، مما يؤدي إلى إنشاء جزيء وظيفي أساسي l l App. بروتين كيناز: الوحدة الفرعية C مسؤولة عن النشاط الأنزيمي، والوحدة الفرعية R مسؤولة عن التنظيم. في البروتينات المتجانسة، الوحدات الفرعية هي نفسها
البنية الرباعية للبروتين l يمكن أن تكون الوحدات الفرعية التي تنشئ البنية الرباعية للبروتين مختلفة تمامًا عن وظائفها الفعلية - البروتينات غير المتجانسة، والتي تتيح لك الجمع بين عدد من الوظائف المترابطة في هيكل واحد، مما يؤدي إلى إنشاء جزيء وظيفي أساسي l l App. بروتين كيناز: الوحدة الفرعية C مسؤولة عن النشاط الأنزيمي، والوحدة الفرعية R مسؤولة عن التنظيم. في البروتينات المتجانسة، الوحدات الفرعية هي نفسها
 البنية الرباعية للبروتين l اتصالات Intersubunit - تم تطوير نظام التفاعلات غير التساهمية بالفعل. البنية الرباعية أقل وأقل ثالثية، مما يعني أن هناك عددًا أقل من الاتصالات الكارهة للماء.
البنية الرباعية للبروتين l اتصالات Intersubunit - تم تطوير نظام التفاعلات غير التساهمية بالفعل. البنية الرباعية أقل وأقل ثالثية، مما يعني أن هناك عددًا أقل من الاتصالات الكارهة للماء.

 وظائف الهيكل الربعي 1. الجمع بين العديد من الوظائف المترابطة في هيكل واحد 2. الوظيفة المعمارية 1. تكوين هياكل واسعة في تكوينات قابلة للطي لضمان قدرة محددة ووظيفية للبروتين (الفيريتين) 2. ملخص التفاعلات الإنزيمية اللاحقة 3. التكوين وظيفية
وظائف الهيكل الربعي 1. الجمع بين العديد من الوظائف المترابطة في هيكل واحد 2. الوظيفة المعمارية 1. تكوين هياكل واسعة في تكوينات قابلة للطي لضمان قدرة محددة ووظيفية للبروتين (الفيريتين) 2. ملخص التفاعلات الإنزيمية اللاحقة 3. التكوين وظيفية
 وظائف البنية الربعية 3. ضمان تفاعلات متعددة للبروتين مع الهياكل الممتدة بروتينات ربط الحمض النووي - dimers (؟) 4. الوظيفة التنظيمية. ينتقل التأثير (الضرر الذي يلحق بالبنية الثلاثية عند التفاعل مع الركيزة) من وحدة فرعية إلى أخرى، مما يؤدي إلى استعادة جميع الهياكل الرباعية
وظائف البنية الربعية 3. ضمان تفاعلات متعددة للبروتين مع الهياكل الممتدة بروتينات ربط الحمض النووي - dimers (؟) 4. الوظيفة التنظيمية. ينتقل التأثير (الضرر الذي يلحق بالبنية الثلاثية عند التفاعل مع الركيزة) من وحدة فرعية إلى أخرى، مما يؤدي إلى استعادة جميع الهياكل الرباعية


 1. يتميز بروتين الجلد الفردي ببنية فريدة من نوعها، مما يضمن تفرد وظيفته. 2. لذلك، يمكن أن يكون فهم بنية البروتينات المختلفة هو المفتاح لفهم طبيعة الأنظمة الحية، وبالتالي، من الواضح، بشكل أساسي
1. يتميز بروتين الجلد الفردي ببنية فريدة من نوعها، مما يضمن تفرد وظيفته. 2. لذلك، يمكن أن يكون فهم بنية البروتينات المختلفة هو المفتاح لفهم طبيعة الأنظمة الحية، وبالتالي، من الواضح، بشكل أساسي
 أدب Berezov T. T.، Korovkin B. F. الكيمياء البيولوجية. - م: الطب، 1983 ميتزلر د. الكيمياء الحيوية. التفاعلات الكيميائية في الخلايا الحية. في 3 مجلدات - مجلدات T. 1، 2. - م: سفيت، 1980 أوفتشينيكوف يو أ. الكيمياء العضوية الحيوية. - م: بروسفيتنيتستفو، 1987 أساسيات الكيمياء الحيوية / إد. أ. أنيسيموفا. - م: فيشا شكولا، 1986 ريس إي، سترينبيرج إم. من الخلايا إلى الذرات. مقدمة مصورة للبيولوجيا الجزيئية. - م: سفيت، 1988 ستيبانوف ف. م. البيولوجيا الجزيئية. هيكل ووظيفة البروتينات. - م: مدرسة فيشا، 1996 بيليبوفيتش يو ب. أساسيات الكيمياء الحيوية. - م: مدرسة فيشا، 1993
أدب Berezov T. T.، Korovkin B. F. الكيمياء البيولوجية. - م: الطب، 1983 ميتزلر د. الكيمياء الحيوية. التفاعلات الكيميائية في الخلايا الحية. في 3 مجلدات - مجلدات T. 1، 2. - م: سفيت، 1980 أوفتشينيكوف يو أ. الكيمياء العضوية الحيوية. - م: بروسفيتنيتستفو، 1987 أساسيات الكيمياء الحيوية / إد. أ. أنيسيموفا. - م: فيشا شكولا، 1986 ريس إي، سترينبيرج إم. من الخلايا إلى الذرات. مقدمة مصورة للبيولوجيا الجزيئية. - م: سفيت، 1988 ستيبانوف ف. م. البيولوجيا الجزيئية. هيكل ووظيفة البروتينات. - م: مدرسة فيشا، 1996 بيليبوفيتش يو ب. أساسيات الكيمياء الحيوية. - م: مدرسة فيشا، 1993


