โครงสร้างของโปรตีนชนิดหนึ่งถูกกำหนดโดยกลุ่มยีน โปรตีน: โครงสร้างโปรตีนปฐมภูมิ, รูปแบบองค์ประกอบไตรเปปไทด์
บิลกิ- สารประกอบอินทรีย์โมเลกุลสูงที่เกิดจากกรดอะมิโนส่วนเกิน
ยู โกดังโปรตีนได้แก่ถ่านหิน น้ำ ไนโตรเจน คิเซน เซอร์กา โปรตีนบางชนิดก่อตัวเป็นสารเชิงซ้อนกับโมเลกุลอื่นๆ เช่น ฟอสฟอรัส เหล็ก สังกะสี และทองแดง
โปรตีนมีน้ำหนักโมเลกุลสูง: อัลบูมินไข่ - 36,000, เฮโมโกลบิน - 152,000, ไมโอซิน - 500,000 สำหรับการเปรียบเทียบ: น้ำหนักโมเลกุลของแอลกอฮอล์ - 46, กรดโอติก - 60, เบนซิน - 78
คลังเก็บกรดอะมิโนของโปรตีน
บิลกิ- โพลีเมอร์ที่ไม่ใช่คาบ เช่น โมโนเมอร์ กรดอะมิโน. กรดα-อะมิโนมี 20 ชนิดที่เรียกว่าโปรตีนโมโนเมอร์ แม้ว่าจะมีการระบุมากกว่า 170 ชนิดในเซลล์และเนื้อเยื่อก็ตาม
สิ่งสำคัญคือต้องทราบว่ากรดอะมิโนสามารถสังเคราะห์ได้ในร่างกายมนุษย์และสิ่งมีชีวิตอื่น ๆ โดยแบ่งออกเป็น: กรดอะมิโนทดแทน- สามารถสังเคราะห์ได้ กรดอะมิโนที่จำเป็น- ไม่สามารถสังเคราะห์ได้ กรดอะมิโนที่จำเป็นจะพบได้ในร่างกายโดยตรงจากร่างกาย พืชสังเคราะห์กรดอะมิโนทุกชนิด
เก็บไว้ในที่เก็บกรดอะมิโน โปรตีนกำลังเต็มที่- พ่นกรดอะมิโนทั้งชุด ด้อยกว่า- กรดอะมิโนทั้งหมดมีอยู่ในสต็อกทุกวัน เนื่องจากโปรตีนประกอบด้วยกรดอะมิโนจึงถูกเรียกว่า ฉันเสียใจ. เมื่อโปรตีนเพิ่มส่วนประกอบที่ไม่ใช่กรดอะมิโน (กลุ่มเทียม) ลงในกรดอะมิโน พวกมันจะถูกเรียกว่า พับเก็บได้. กลุ่มเทียมสามารถแสดงได้ด้วยโลหะ (เมตาโลโปรตีน), คาร์โบไฮเดรต (ไกลโคโปรตีน), ไขมัน (ไลโปโปรตีน), กรดนิวคลีอิก (นิวคลีโอโปรตีน)
หนวด กรดอะมิโนจะแก้แค้น: 1) หมู่คาร์บอกซิล (-COOH); 2) หมู่อะมิโน (-NH 2); 3) อนุมูลหรือกลุ่ม R (โมเลกุล reshta) Budova Radical แตกต่างกันไปตามกรดอะมิโนประเภทต่างๆ ขึ้นอยู่กับจำนวนหมู่อะมิโนและหมู่คาร์บอกซิลที่เข้าสู่คลังกรดอะมิโน พวกมันจะถูกแบ่งออกเป็น: กรดอะมิโนที่เป็นกลางมีกลุ่มคาร์บอกซิลหนึ่งกลุ่มและกลุ่มอะมิโนหนึ่งกลุ่ม กรดอะมิโนพื้นฐานมีกลุ่มอะมิโนมากกว่าหนึ่งกลุ่ม กรดอะมิโนที่เป็นกรดมากกว่าหนึ่งหมู่คาร์บอกซิลคืออะไร?
กรดอะมิโน สารกึ่งตัวนำแบบแอมโฟเทอริกดังนั้นในอาหาร กลิ่นเหม็นอาจเกิดจากทั้งกรดและเบส ในน้ำหลากหลายชนิด กรดอะมิโนจะปรากฏในรูปแบบไอออนิกที่แตกต่างกัน
ลิงค์เปปไทด์
เปปติดี- สารประกอบอินทรีย์ที่เกิดขึ้นจากกรดอะมิโนส่วนเกินที่เชื่อมต่อกันด้วยพันธะเปปไทด์
การปลดปล่อยเปปไทด์เกิดขึ้นตามปฏิกิริยาการควบแน่นของกรดอะมิโน เมื่อหมู่อะมิโนของกรดอะมิโนตัวหนึ่งทำปฏิกิริยากับหมู่คาร์บอกซิลของอีกกลุ่มหนึ่ง จะเกิดการเชื่อมโยงระหว่างไนโตรเจนและคาร์โบไฮเดรตแบบโควาเลนต์ ซึ่งเรียกว่า เปปไทด์. สิ่งสำคัญคือต้องแยกปริมาณกรดอะมิโนส่วนเกินที่ไปสะสมในเปปไทด์ออกจากกัน ดิเปปติดี, ทริปเปปติดี, เตตราเปปติดีฯลฯ การก่อตัวของพันธะเปปไทด์สามารถเกิดซ้ำได้หลายวิธี นำสิ่งนี้มาสู่ความกระจ่าง โพลีเปปไทด์. ที่ปลายด้านหนึ่งของเปปไทด์จะมีหมู่อะมิโนอิสระ (เรียกว่า ปลายเอ็น) และปลายอีกด้านหนึ่งมีหมู่คาร์บอกซิลอิสระ (เรียกว่า ปลายซี)

การจัดเรียงโมเลกุลโปรตีนอย่างง่าย
หน้าที่เฉพาะของโปรตีนขึ้นอยู่กับการกำหนดค่าเชิงพื้นที่ของโมเลกุล นอกจากนี้ เซลล์จะตัดโปรตีนในรูปแบบเปิดอย่างมีพลังและมองไม่เห็นซึ่งดูเหมือนหอก ดังนั้นหอกโพลีเปปไทด์จึงมีการก่อตัวและการบวมของโครงสร้างและโครงสร้างเล็กน้อย ดู 4 ภูมิภาค การจัดระเบียบโปรตีนที่กว้างขวาง.
โครงสร้างโปรตีนปฐมภูมิ- ลำดับการละลายของกรดอะมิโนส่วนเกินจากโพลีเปปไทด์มีดหมอเพื่อสร้างโมเลกุลโปรตีน พันธะระหว่างกรดอะมิโนคือเปปไทด์

เนื่องจากโมเลกุลโปรตีนประกอบด้วยกรดอะมิโนส่วนเกินเพียง 10 ตัว ดังนั้นโมเลกุลโปรตีนที่เป็นไปได้ในทางทฤษฎีซึ่งแปรผันตามลำดับการเติมกรดอะมิโนคือ 10 20 ด้วยกรดอะมิโน 20 ชนิด คุณสามารถรวมพวกมันเข้าด้วยกันเป็นส่วนผสมที่แตกต่างกันมากยิ่งขึ้น มีการระบุโปรตีนที่แตกต่างกันประมาณหมื่นชนิดในมนุษย์ ซึ่งแตกต่างกันไปในประเภทหนึ่งไปยังอีกประเภทหนึ่ง เช่นเดียวกับโปรตีนของสิ่งมีชีวิตอื่น ๆ
โครงสร้างปฐมภูมิของโมเลกุลโปรตีนบ่งบอกถึงพลังของโมเลกุลโปรตีนและโครงสร้างที่กว้างขวางของมัน การแทนที่กรดอะมิโนตัวหนึ่งด้วยอีกตัวหนึ่งในสายโซ่โพลีเปปไทด์ทำให้เกิดการเปลี่ยนแปลงในด้านอำนาจและหน้าที่ของโปรตีน ตัวอย่างเช่นการแทนที่กรดอะมิโนกลูตามิกในหน่วยβ-subunit ของเฮโมโกลบินด้วยวาลีนนำไปสู่ความจริงที่ว่าโมเลกุลของฮีโมโกลบินโดยรวมไม่สามารถสูญเสียหน้าที่หลักได้นั่นคือการขนส่งกรด ในกรณีเช่นนี้ ผู้คนจะมีอาการป่วย - โรคโลหิตจางชนิดเคียว
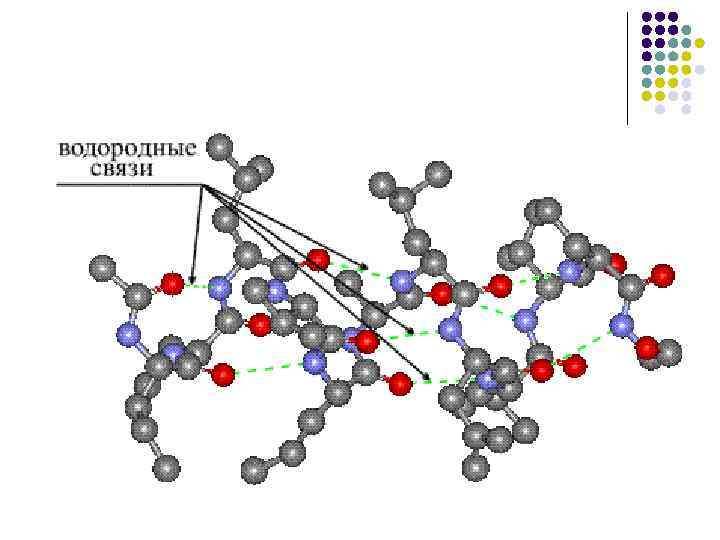
โครงสร้างรอง- กล่องเสียงของมีดหมอโพลีเปปไทด์ถูกจัดเรียงเป็นเกลียว (ดูเหมือนสปริงขยายออก) ขดลวดของเกลียวนั้นเกิดจากพันธะน้ำที่พันกันระหว่างหมู่คาร์บอกซิลและหมู่อะมิโน กลุ่ม CO และ NH เกือบทั้งหมดมีส่วนร่วมในการสร้างการเชื่อมต่อทางน้ำ กลิ่นเหม็นจะอ่อนกว่าเปปไทด์ แต่เมื่อทำซ้ำแล้วจะเต็มไปด้วยก๊าซซึ่งทำให้การเปลี่ยนแปลงเหล่านี้คงอยู่และรุนแรง ในระดับโครงสร้างทุติยภูมิมีโปรตีน: ไฟโบรอิน (รอยประสาน, ไหม), เคราติน (ผม, เล็บ), คอลลาเจน (เส้นเอ็น)



โครงสร้างเทรติน- อีตัวของครึ่ง -petd Lancyugiv ที่ Globuli, viniki veneolikenny ของ vinikenni ของ ninesty Zv'yazkiv (vodnevikh, disulfіdni) คือความเสียหายของ girdrophic mira ร่วมกันโดยอนุมูลของกรดอะมิโน บทบาทหลักของโครงสร้างตติยภูมิที่ส่องสว่างนั้นเล่นโดยปฏิกิริยาที่ชอบน้ำและไม่ชอบน้ำ ในโมเลกุลที่เป็นน้ำ อนุมูลที่ไม่ชอบน้ำมีแนวโน้มที่จะมาบรรจบกันในน้ำ โดยรวมตัวกันที่ตรงกลางของทรงกลม ในขณะที่อนุมูลที่ชอบน้ำซึ่งเป็นผลมาจากการให้ความชุ่มชื้น (อันตรกิริยากับไดโพลของน้ำ) มีแนวโน้มที่จะเกาะอยู่บนพื้นผิวของโมเลกุล ในโปรตีนบางชนิด โครงสร้างตติยภูมิจะเสถียรโดยพันธะโควาเลนต์ไดซัลไฟด์ที่เกิดขึ้นระหว่างอะตอมคาร์บอนของซิสเทอีนส่วนเกินสองตัว ในระดับโครงสร้างตติยภูมิจะมีเอนไซม์ แอนติบอดี และฮอร์โมนบางชนิด

โครงสร้างควอเตอร์นารีและในโปรตีนพับหมายถึงโมเลกุลที่ก่อตัวเป็นสองกลมขึ้นไป หน่วยย่อยจะรวมกันอยู่ในโมเลกุลผ่านปฏิกิริยาระหว่างไอออนิก ไม่ชอบน้ำ และไฟฟ้าสถิต บางครั้งเมื่อมีการสร้างโครงสร้างควอเทอร์นารี พันธะไดซัลไฟด์จะปรากฏขึ้นระหว่างหน่วยย่อย โปรตีนที่ใหญ่ที่สุดที่มีโครงสร้างหนึ่งในสี่คือ เฮโมโกลบิน. ประกอบด้วยหน่วยย่อย α สองหน่วย (กรดอะมิโนส่วนเกิน 141 หน่วย) และหน่วยย่อย β สองหน่วย (กรดอะมิโนส่วนเกิน 146 หน่วย) โมเลกุลของฮีมมีความเกี่ยวข้องกับหน่วยย่อยของผิวหนังซึ่งมีหน้าที่รับผิดชอบในการปลดปล่อย
ไม่ว่าด้วยเหตุผลใดก็ตาม หากโครงสร้างโปรตีนเบี่ยงเบนไปจากปกติ โปรตีนจะไม่สามารถคงหน้าที่ของมันไว้ได้ ตัวอย่างเช่น สาเหตุของ "เรื่องวัว" (โรคไข้สมองอักเสบสปองจิฟอร์ม) คือโครงสร้างที่ผิดปกติของพรีออน - โปรตีนบนพื้นผิวของเซลล์ประสาท
พลังแห่งกระรอก
การเก็บกรดอะมิโน โครงสร้างของโมเลกุลโปรตีนหมายถึง พลัง. โปรตีนได้รับความเป็นกรดพื้นฐานซึ่งถูกกำหนดโดยอนุมูลของกรดอะมิโน: ยิ่งโปรตีนมีกรดที่เป็นกรดมากเท่าใด การแสดงออกของความเป็นกรดก็จะยิ่งสดใสมากขึ้นเท่านั้น คุณค่าของการให้และการมา H+ หมายถึง พลังบัฟเฟอร์ของโปรตีน; บัฟเฟอร์ที่สำคัญที่สุดอย่างหนึ่งคือฮีโมโกลบินในเซลล์เม็ดเลือดแดง ซึ่งช่วยรักษาค่า pH ของเลือดให้อยู่ในระดับคงที่ - โปรตีนที่แยกจากกัน (ไฟบริโนเจน) และโปรตีนที่ไม่แยกจากกันซึ่งมีส่วนช่วยในการทำงานทางกล (ไฟบริน, เคราติน, คอลลาเจน) มีโปรตีนที่ทำงานอยู่ในรูปแบบทางเคมี (เอนไซม์) และมีโปรตีนที่ไม่ใช้งานทางเคมี ทนต่อการไหลบ่าเข้ามาของจิตใจที่แตกต่างจากโลกภายนอก และไม่เสถียรอย่างยิ่ง
ปัจจัยภายนอก (ความร้อน, การสัมผัสรังสีอัลตราไวโอเลต, โลหะสำคัญและเกลือ, การเปลี่ยนแปลง pH, การแผ่รังสี, มลภาวะทางน้ำ)
อาจทำให้เกิดความเสียหายต่อการจัดโครงสร้างของโมเลกุลโปรตีนได้ เรียกว่ากระบวนการสูญเสียโครงสร้างเล็กน้อยซึ่งเป็นโมเลกุลโปรตีนที่โดดเด่น การเสียสภาพ. สาเหตุของการเสียสภาพคือการแตกของเอ็นที่ทำให้โครงสร้างโปรตีนมีความเสถียร เส้นเอ็นที่อ่อนแอที่สุดจะฉีกขาดในไต และเมื่อจิตใจแข็งแรงขึ้น เส้นเอ็นที่แข็งแรงก็จะฉีกขาด จากนั้นจึงมีการพัฒนาโครงสร้างควอเทอร์นารีจากนั้นระดับอุดมศึกษาและระดับทุติยภูมิ การเปลี่ยนโครงสร้างเชิงพื้นที่ทำให้เกิดการเปลี่ยนแปลงในพลังของโปรตีน และเป็นผลให้โปรตีนไม่สามารถสูญเสียการควบคุมการทำงานทางชีวภาพของมันได้ เนื่องจากการสูญเสียสภาพไม่ได้มาพร้อมกับการทำลายโครงสร้างหลักจึงอาจเกิดขึ้นได้ มนุษย์หมาป่าเมื่อมีการต่ออายุโครงสร้างโปรตีนพลังงานด้วยตนเอง การสูญเสียสภาพดังกล่าวได้รับการยอมรับ ตัวอย่างเช่น โดยโปรตีนของตัวรับเมมเบรน เรียกว่ากระบวนการต่ออายุโครงสร้างโปรตีนหลังจากการทำให้เสียสภาพ การคืนสภาพ. หากเป็นไปไม่ได้ที่จะต่ออายุการกำหนดค่าเชิงพื้นที่ของโปรตีนก็จะเรียกว่าการสูญเสียสภาพธรรมชาติ เอาคืนไม่ได้.
หน้าที่ของโปรตีน
| การทำงาน | กรุณาให้คำอธิบาย |
|---|---|
| บูดิเวลนา | โปรตีนมีส่วนร่วมในการก่อตัวของโครงสร้างเซลล์และหลังคลินิก: พวกมันเข้าไปในเยื่อหุ้มเซลล์ (ไลโปโปรตีน, ไกลโคโปรตีน), ผม (เคราติน), เส้นเอ็น (คอลลาเจน) เป็นต้น |
| ขนส่งนา | โปรตีนฮีโมโกลบินในเลือดจะเพิ่มความเป็นกรดและขนส่งจากเลือดไปยังเนื้อเยื่อและอวัยวะทั้งหมด จากนั้นจะส่งก๊าซคาร์บอนไดออกไซด์จากเลือดเหล่านั้น การจัดเก็บเยื่อหุ้มเซลล์ประกอบด้วยโปรตีนพิเศษที่ช่วยให้มั่นใจได้ถึงการถ่ายโอนสารและไอออนบางชนิดจากเซลล์ไปยังแกนนอกและด้านหลังอย่างเข้มงวดและเข้มงวด |
| กฎระเบียบ | ฮอร์โมนโปรตีนมีส่วนร่วมในการควบคุมกระบวนการเผาผลาญคำพูด ตัวอย่างเช่น ฮอร์โมนอินซูลินควบคุมระดับกลูโคสในเลือด ส่งเสริมการสังเคราะห์ไกลโคเจน และเพิ่มการลดไขมันจากคาร์โบไฮเดรต |
| ซาฮิสนา | เมื่อโปรตีนและจุลินทรีย์จากต่างประเทศ (แอนติเจน) แทรกซึมเข้าไปในร่างกาย โปรตีนชนิดพิเศษจะถูกสร้างขึ้น - แอนติบอดีซึ่งจับและขัดผิว ไฟบรินซึ่งสร้างด้วยไฟบริโนเจนช่วยลดการตกเลือด |
| ทวิกูน่า | โปรตีนแอคตินและไมโอซินที่มีอายุสั้นช่วยให้แผลในสัตว์ที่มีเซลล์สมบูรณ์สั้นลง |
| ซิกแนลนา | ที่เยื่อหุ้มผิวของเซลล์มีโมเลกุลโปรตีนที่เปลี่ยนโครงสร้างตติยภูมิเพื่อตอบสนองต่อปัจจัยของตัวกลางภายนอก จึงรับสัญญาณจากตัวกลางภายนอกและส่งคำสั่งไปยังเซลล์. |
| ตุนขึ้น | ในร่างกายของสัตว์โปรตีนตามกฎจะไม่ถูกเก็บไว้ผู้ร้าย: อัลบูมินไข่เคซีนนม อย่างไรก็ตามโปรตีนในร่างกายสามารถจัดเตรียมคำพูดไว้ได้เช่นเมื่อฮีโมโกลบินแตกตัวของเหลวจะไม่ถูกขับออกจากร่างกาย แต่จะถูกเก็บไว้ซึ่งสร้างโปรตีนเชิงซ้อนด้วยเฟอร์ริติน |
| กระฉับกระเฉง | เมื่อโปรตีน 1 กรัมสลายตัวไปจนถึงผลิตภัณฑ์ขั้นสุดท้าย จะมองเห็นได้ 176 กิโลจูล ในตอนแรก โปรตีนจะแตกตัวเป็นกรดอะมิโน จากนั้นจึงกลายเป็นผลิตภัณฑ์ขั้นสุดท้าย เช่น น้ำ คาร์บอนไดออกไซด์ และแอมโมเนีย อย่างไรก็ตาม ในฐานะแหล่งพลังงาน โปรตีนจะถูกดูดซึมก็ต่อเมื่อมีการบริโภคแหล่งอื่น (คาร์โบไฮเดรตและไขมัน) เท่านั้น |
| ตัวเร่งปฏิกิริยา | หน้าที่ที่สำคัญที่สุดอย่างหนึ่งของโปรตีน ให้โปรตีน - เอนไซม์ที่ช่วยเร่งปฏิกิริยาทางชีวเคมีที่เกิดขึ้นในเซลล์ ตัวอย่างเช่นไรบูโลสไบฟอสเฟตคาร์บอกซิเลสเร่งการตรึง 2 ในระหว่างการสังเคราะห์ด้วยแสง |
เฟอร์เมนติ
เฟอร์เมนติ, หรือ เอนไซม์, - โปรตีนประเภทพิเศษที่เป็นตัวเร่งปฏิกิริยาทางชีวภาพ เนื่องจากเอนไซม์ ปฏิกิริยาทางชีวเคมีจึงมีความลื่นไหลสูง ความเร็วของปฏิกิริยาเอนไซม์เป็นหมื่นครั้ง (และเป็นล้านครั้ง) เกิดจากความเร็วของปฏิกิริยาที่เกิดขึ้นจากการมีส่วนร่วมของตัวเร่งปฏิกิริยาอนินทรีย์ เรียกว่า Rechovina ซึ่งเป็นสิ่งที่เอนไซม์ให้ผล วัสดุพิมพ์.
เอนไซม์ - โปรตีนทรงกลม ลักษณะเฉพาะของบูโดวีเอนไซม์สามารถแบ่งออกเป็นสองกลุ่ม: แบบง่ายและซับซ้อน ฉันขอโทษที่หมักด้วยโปรตีนที่ง่ายที่สุดแล้ว เกิดจากกรดอะมิโนเป็นหลัก เอนไซม์พับєพับผ้าขาวแล้ว ก่อนถึงโกดัง นอกจากส่วนโปรตีนแล้วยังมีกลุ่มที่มีลักษณะไม่ใช่โปรตีนอีกด้วย ปัจจัยร่วม. ในเอนไซม์บางชนิด วิตามินจะทำหน้าที่เป็นโคแฟคเตอร์ โมเลกุลของเอนไซม์มีส่วนพิเศษที่เรียกว่าแอคทีฟเซ็นเตอร์ ศูนย์ที่ใช้งานอยู่- ส่วนเล็กๆ ของเอนไซม์ (ส่วนเกินของกรดอะมิโนสามถึงสิบสอง) ซึ่งจับกับสารตั้งต้นหรือสารตั้งต้นเข้ากับสารเชิงซ้อนของเอนไซม์-สารตั้งต้น หลังจากเสร็จสิ้นการทำปฏิกิริยา สารเชิงซ้อนของเอนไซม์-ซับสเตรตจะแตกตัวออกเป็นเอนไซม์และผลิตภัณฑ์ที่ทำปฏิกิริยา เอนไซม์ที่ใช้งานจริง (แอคทีฟครีม) ศูนย์อัลโลสเตอริก- ส่วนที่ใช้ควบคุมสภาพคล่องและเอนไซม์ ( เอนไซม์อัลโลสเตอริก).

ปฏิกิริยาของการเร่งปฏิกิริยาด้วยเอนไซม์มีลักษณะเฉพาะคือ 1) ประสิทธิภาพสูง 2) การคัดเลือกอย่างเข้มงวดและความตรงของการออกฤทธิ์ 3) ความจำเพาะของสารตั้งต้น 4) การควบคุมที่ละเอียดและแม่นยำ ความจำเพาะของสารตั้งต้นและปฏิกิริยาของปฏิกิริยาของการเร่งปฏิกิริยาด้วยเอนไซม์อธิบายได้จากสมมติฐานของ E. Fisher (1890) และ D. Koshland (1959)
อี. ฟิชเชอร์ (สมมติฐานการล็อคกุญแจ)สมมติว่าการกำหนดค่าที่กว้างของแอคทีฟเซ็นเตอร์ของเอนไซม์และซับสเตรตนั้นเหมือนกันทุกประการ สารตั้งต้นเหมือนกับ "กุญแจ" เอนไซม์คือ "ล็อค"
D. Koshland (สมมติฐานถุงมือมือ)สมมติว่าความคล้ายคลึงกันของโครงสร้างระหว่างสารตั้งต้นและจุดศูนย์กลางแอคทีฟของเอนไซม์นั้นถูกสร้างขึ้นในช่วงเวลาที่มีปฏิสัมพันธ์กันทีละรายการเท่านั้น สมมติฐานนี้เรียกอีกอย่างว่า สมมติฐานของสายพันธุ์ที่ถูกชักนำ.
ความลื่นไหลของปฏิกิริยาของเอนไซม์ขึ้นอยู่กับ: 1) อุณหภูมิ; 2) ความเข้มข้นของเอนไซม์ 3) ความเข้มข้นของสารตั้งต้น; 4) ค่า pH ควรสังเกตว่าชิ้นส่วนของเอนไซม์นั้นเป็นโปรตีนดังนั้นกิจกรรมของพวกมันจะสูงที่สุดในจิตใจปกติทางสรีรวิทยา
เอนไซม์ส่วนใหญ่สามารถแปรรูปได้ที่อุณหภูมิตั้งแต่ 0 ถึง 40 °C เท่านั้น ในบริเวณเหล่านี้ ความลื่นไหลของปฏิกิริยาจะเพิ่มขึ้นประมาณ 2 เท่าเมื่ออุณหภูมิผิวหนังเพิ่มขึ้นถึง 10 °C ที่อุณหภูมิสูงกว่า 40 ° C โปรตีนจะเกิดการเสื่อมสภาพและกิจกรรมของเอนไซม์จะลดลง ที่อุณหภูมิใกล้กับจุดเยือกแข็ง เอนไซม์จะถูกปิดใช้งาน
เมื่อปริมาณสารตั้งต้นเพิ่มขึ้น ความลื่นไหลของปฏิกิริยาของเอนไซม์จะเพิ่มขึ้นจนกระทั่งจำนวนโมเลกุลในสารตั้งต้นจะเท่ากับจำนวนโมเลกุลในเอนไซม์ เมื่อปริมาณของสารตั้งต้นเพิ่มขึ้นอีก สภาพคล่องจะไม่เพิ่มขึ้นตราบใดที่ศูนย์กลางของเอนไซม์ยังอิ่มตัวอยู่ ความเข้มข้นของเอนไซม์ที่สูงขึ้นนำไปสู่กิจกรรมการเร่งปฏิกิริยาที่เพิ่มขึ้น เนื่องจากในหนึ่งชั่วโมงของปฏิกิริยา โมเลกุลจำนวนมากจะสัมผัสกับสารตั้งต้น

สำหรับเอนไซม์ผิวหนัง มีค่า pH ที่เหมาะสมซึ่งจะแสดงกิจกรรมสูงสุด (เปปซิน – 2.0, เมือกอะไมเลส – 6.8, ไลเปสใต้ผิวหนัง – 9.0) ที่ค่า pH สูงหรือต่ำ กิจกรรมของเอนไซม์จะลดลง ในกรณีที่มีการเปลี่ยนแปลง pH อย่างรุนแรง เอนไซม์จะเสื่อมสภาพ
ความลื่นไหลของเอนไซม์อัลโลสเตอริกถูกควบคุมโดยสารที่ไปถึงศูนย์กลางอัลโลสเตอริก เนื่องจากคำเหล่านี้เร่งปฏิกิริยาจึงเรียกว่าเหม็น ตัวกระตุ้น, yakscho ถึง galm สารยับยั้ง.
การจำแนกประเภทของเอนไซม์
ขึ้นอยู่กับประเภทของปฏิกิริยาเคมีที่เร่งปฏิกิริยา เอนไซม์แบ่งออกเป็น 6 ประเภท:
- ออกซิรีดักเตส(การถ่ายโอนอะตอมจากน้ำ กรด หรืออิเล็กตรอนจากสารหนึ่งไปยังอีกสารหนึ่ง - ดีไฮโดรจีเนส)
- การโอน(การถ่ายโอนของกลุ่มเมทิล, อะซิล, ฟอสเฟตหรืออะมิโนจากโมเลกุลหนึ่งไปยังอีกโมเลกุลหนึ่ง - ทรานซามิเนส)
- ไฮโดรเลส(ปฏิกิริยาไฮโดรไลซิสซึ่งผลิตภัณฑ์ทั้งสองถูกสังเคราะห์จากสารตั้งต้น - อะไมเลส, ไลเปส)
- ลิอาซี(การเติมแบบไม่ไฮโดรไลติกให้กับสารตั้งต้นหรือการแยกออกจากอะตอมกลุ่มใหม่ในระหว่างที่พันธะ C-C, C-N, C-O, C-S decarboxylase สามารถแตกได้)
- ไอโซเมอเรส(perebudova ในโมเลกุล - ไอโซเมอเรส)
- ลิกาซี(การก่อตัวของสองโมเลกุลอันเป็นผลมาจากการก่อตัวของพันธะ C-C, C-N, C-O, C-S - synthetase)
จำแนกภาพวาดของคุณออกเป็นคลาสย่อยและคลาสย่อย ในการจำแนกระหว่างประเทศที่มีอยู่ เอนไซม์ผิวหนังมีรหัสเฉพาะที่ประกอบด้วยตัวเลขสี่ตัวคั่นด้วยจุด หมายเลขแรกคือคลาส อีกอันคือคลาสย่อย หมายเลขที่สามคือคลาสย่อย หมายเลขที่สี่คือหมายเลขซีเรียลของเอนไซม์ในคลาสย่อยนั้น ตัวอย่างเช่น รหัสอาร์จิเนสคือ 3.5.3.1
ไปที่ การบรรยายครั้งที่ 2“หน้าที่ของคาร์โบไฮเดรตและไขมันคืออะไร”
ไปที่ การบรรยายครั้งที่ 4“หน้าที่ของกรดนิวคลีอิก ATP”
การสังเคราะห์โปรตีน
1. กำหนดโครงสร้างของโปรตีนหนึ่งชนิด:
1) กลุ่มของยีน 2) หนึ่งจีโนม
3) หนึ่งโมเลกุล DNA 4) จำนวนทั้งหมดของยีนในสิ่งมีชีวิต
2. ยีนเข้ารหัสข้อมูลเกี่ยวกับลำดับของโมโนเมอร์ในโมเลกุล:
1) tRNA 2) AA 3) ไกลโคเจน 4) ดีเอ็นเอ
3. แฝดสามเรียกว่าแอนติโคดอน:
1) ดีเอ็นเอ 2) ที-อาร์เอ็นเอ 3) ไอ-อาร์เอ็นเอ 4) อาร์-อาร์เอ็นเอ
4. การแลกเปลี่ยนพลาสติกพัฒนาที่สำคัญเมื่อมีปฏิกิริยา:
1) การสลายตัวของสุนทรพจน์อินทรีย์ 2) การสลายตัวของสุนทรพจน์อนินทรีย์
3) การสังเคราะห์สารอินทรีย์ 4) การสังเคราะห์สารอนินทรีย์
5. การสังเคราะห์โปรตีนในเซลล์โปรคาริโอตเกิดขึ้น:
1) บนไรโบโซมในนิวเคลียส 2) บนไรโบโซมในไซโตพลาสซึม 3) ที่บริเวณเซลล์
4) บนพื้นผิวด้านนอกของเมมเบรนไซโตพลาสซึม
6. กระบวนการออกอากาศเริ่มต้นขึ้น:
1) ในไซโตพลาสซึม 2) ในนิวเคลียส 3) ในไมโตคอนเดรีย
4) บนเยื่อหุ้มของเยื่อหุ้มเอนโดพลาสซึมสั้น
7. บนเยื่อหุ้มของเยื่อเอนโดพลาสซึมแบบเม็ดการสังเคราะห์เกิดขึ้น:
1) เอทีพี; 2) ในคาร์โบไฮเดรต; 3) ไขมัน; 4) โปรตีน
8. รหัสแฝดหนึ่งรหัส:
1. AK หนึ่งอัน 2 สัญญาณของร่างกายหนึ่งอัน 3. เฝือก AK
9. การสังเคราะห์โปรตีนเสร็จสมบูรณ์ในขณะนี้
1. การจดจำโคดอนโดยแอนติโคดอน 2. ลักษณะของ “เครื่องหมายแบ่ง” บนไรโบโซม
3. ถ่ายโอน i-RNA ไปยังไรโบโซม
10. กระบวนการอ่านข้อมูลจากโมเลกุล DNA
1. การแปล 2. การถอดความ 3. การเปลี่ยนแปลง
11.พลังของโปรตีนถูกกำหนด...
1. โครงสร้างรองของโปรตีน 2. โครงสร้างปฐมภูมิของโปรตีน
3.โครงสร้างโปรตีนระดับตติยภูมิ
12. กระบวนการที่แอนติโคดอนได้รับการยอมรับว่าเป็นโคดอนบน i-RNA
13. ขั้นตอนของการสังเคราะห์โปรตีน
1.การถอดความ การแปล 2.การเปลี่ยนแปลง การแปล
3. การย้ายองค์กร การถอดความ
14. Anticodon t-RNA ประกอบด้วยนิวคลีโอไทด์ UCG DNA Triplet ใดที่ส่งเสริมคุณ?
1.UUG 2. TTC 3. บสย
15. จำนวน t-RNA ที่มีส่วนร่วมในการแปลเป็นจำนวนดั้งเดิม:
1. รหัสของ i-RNA ที่เข้ารหัสกรดอะมิโน 2. โมเลกุล i-RNA
3 ยีนที่รวมอยู่ก่อนโมเลกุล DNA 4. โปรตีนที่สังเคราะห์บนไรโบโซม
16. กำหนดลำดับของนิวคลีโอไทด์ของ i-RNA ในระหว่างการถอดรหัสจากหนึ่งในสาย DNA: A-G-T-C-G
1) คุณ 2) ก 3) ค 4) ก 5) ค
17. ในระหว่างการจำลองโมเลกุล DNA จะมีสิ่งต่อไปนี้เกิดขึ้น:
1) ด้ายที่แตกออกเป็นชิ้นเล็ก ๆ ของโมเลกุลลูกสาว
2) โมเลกุลที่ประกอบด้วยสาย DNA ใหม่สองเส้น
3) โมเลกุล ครึ่งหนึ่งประกอบด้วยสาย iRNA
4) โมเลกุลลูกสาวที่เกิดจาก DNA สายเก่าและสายใหม่หนึ่งสาย
18. เทมเพลตสำหรับการสังเคราะห์โมเลกุล iRNA ระหว่างการถอดรหัสคือ:
1) โมเลกุล DNA ทั้งหมด 2) พื้นผิวของโมเลกุล DNA อันใดอันหนึ่ง
3) โครงเรื่องของหนึ่งใน DNA ของ Lancs
4) ในบางประเภทหนึ่งในโมเลกุล DNA ในประเภทอื่น ๆ – โมเลกุล DNA ทั้งหมด
19. กระบวนการมุดตัวของโมเลกุล DNA
1. การจำลองแบบ 2. การซ่อมแซม
3. การกลับชาติมาเกิด
20. ในระหว่างการสังเคราะห์โปรตีนในเซลล์ พลังงาน ATP:
1)ใช้จ่าย 2)ตุน
3) ไม่ได้ใช้และไม่เห็น
21. ในเซลล์ร่างกายของสิ่งมีชีวิตเซลล์สมบูรณ์:
1) ชุดของยีนและโปรตีนที่แตกต่างกัน 2) ชุดของยีนและโปรตีนชุดใหม่
3) ยีนชุดใหม่หรือโปรตีนชุดอื่น
4) โปรตีนชุดใหม่หรือยีนชุดอื่น
22.. DNA แฝดสามอันประกอบด้วยข้อมูลเกี่ยวกับ:
1) ลำดับของกรดอะมิโนในโมเลกุลโปรตีน
2) สัญญาณของร่างกาย 3) กรดอะมิโนในโมเลกุลโปรตีนสังเคราะห์
4) การจัดเก็บโมเลกุล RNA
23. กระบวนการใดที่ไม่มีอยู่ในเซลล์ของฟังก์ชันใด ๆ :
1) การสังเคราะห์โปรตีน 2) เมแทบอลิซึมของโปรตีน 3) ไมโทซิส 4) ไมโอซิส
24. แนวคิดของ “การถอดความ” หมายถึงกระบวนการ:
1) การมุดตัวของ DNA 2) การสังเคราะห์ i-RNA บน DNA
3) การเปลี่ยน i-RNA ไปเป็นไรโบโซม 4) การสร้างโมเลกุลโปรตีนบนโพลีโซม
25. ส่วนหนึ่งของโมเลกุล DNA ที่บรรจุข้อมูลเกี่ยวกับโมเลกุลโปรตีนหนึ่งโมเลกุล:
1) ยีน 2) ฟีโนไทป์ 3) จีโนม 4) จีโนไทป์
26. การถอดความในยูคาริโอตเกิดขึ้นใน:
1) ไซโตพลาสซึม 2) เยื่อหุ้มเอนโดพลาสซึม 3) ไลโซโซม 4) นิวเคลียส
27. การสังเคราะห์โปรตีนได้มาจาก:
1) ตาข่ายเอนโดพลาสซึมแบบละเอียด
2) เรติเคิลเอนโดพลาสมิกเรียบ 3) นิวเคลียส 4) ไลโซโซม
28. กรดอะมิโนตัวหนึ่งมีรหัส:
1) นิวคลีโอไทด์หลายตัว 2) นิวคลีโอไทด์สองตัว
3) หนึ่งนิวคลีโอไทด์ 4) สามนิวคลีโอไทด์
29. นิวคลีโอไทด์ ATC แฝดในโมเลกุล DNA มีโคดอนคล้ายกับโมเลกุล i-RNA:
1) แท็ก 2) UAG 3) UTC 4) TsAU
30. ป้ายแบ่งรหัสพันธุกรรม:
1.เข้ารหัสโปรตีนเพลง 2.กระตุ้นการสังเคราะห์โปรตีน
3.ส่งเสริมการสังเคราะห์โปรตีน
31. กระบวนการสมัครสมาชิกโมเลกุล DNA ด้วยตนเอง
1.การจำลอง 2.การซ่อมแซม 3.การรีแอคชั่น
32. หน้าที่ของ i-RNA ในกระบวนการสังเคราะห์ทางชีวภาพ
1.การบันทึกข้อมูลการบดอัด 2.การขนส่ง AK ไปยังไรโบโซม
3. ส่งข้อมูลให้กับไรโบโซม
33. กระบวนการที่ t-RNA นำกรดอะมิโนไปที่ไรโบโซม
1.การถอดความ 2.การแปล 3.การเปลี่ยนแปลง
34. ไรโบโซม ซึ่งสังเคราะห์โมเลกุลโปรตีนเดียวกัน
1.โครโมโซม 2.โพลีโซม 3.เมกาโครโมโซม
35. กระบวนการที่กรดอะมิโนทำให้โมเลกุลโปรตีนคงตัว
1.การถอดความ 2.การแปล 3.การเปลี่ยนแปลง
36. ก่อนเกิดปฏิกิริยาการสังเคราะห์เมทริกซ์ เรานำ...
1.การจำลองแบบ DNA 2.การถอดความ การแปล 3.ทั้งสองประเภทถูกต้อง
37. DNA triplet หนึ่งอันมีข้อมูลเกี่ยวกับ:
1. ลำดับของกรดอะมิโนในโมเลกุลโปรตีน
2.สถานที่ร้องเพลง AK ในมีดหมอโปรตีน
3. สัญลักษณ์ของสิ่งมีชีวิตเฉพาะ
4. กรดอะมิโนที่รวมอยู่ในโปรตีนมีดหมอ
38. Genya ได้เข้ารหัสข้อมูลเกี่ยวกับ:
1) โครงสร้างของโปรตีน ไขมัน และคาร์โบไฮเดรต 2) โครงสร้างปฐมภูมิของโปรตีน
3) ลำดับของนิวคลีโอไทด์ใน DNA
4) ลำดับของกรดอะมิโนในโมเลกุลโปรตีน 2 โมเลกุลขึ้นไป
39. การสังเคราะห์ mRNA เริ่มต้นด้วย:
1) การแยก DNA ออกเป็นสองสาย 2) อันตรกิริยากับเอนไซม์ RNA polymerase และยีน
3) การสืบทอดของยีน 4) การสลายตัวของยีนไปเป็นนิวคลีโอไทด์
40. มีการถอดความ:
1) ในนิวเคลียส 2) บนไรโบโซม 3) ในไซโตพลาสซึม 4) บนช่อง EPS แบบเรียบ
41. การสังเคราะห์โปรตีนไม่เกิดขึ้นกับไรโบโซมใน:
1) วัณโรค zbudnik 2) bjoli 3) แมลงวันเห็ด 4) แบคทีเรีย
42. ในระหว่างการแปล เมทริกซ์สำหรับการประกอบโปรตีนโพลีเปปไทด์คือ:
1) หนึ่งในสาย DNA 2) หนึ่งในโมเลกุล DNA
3) โมเลกุล iRNA 4) ในบางรูปแบบหนึ่งในสาย DNA ในรูปแบบอื่น ๆ – โมเลกุล iRNA
โครงสร้างหลักของโปรตีนคือโพลีเปปไทด์เชิงเส้นที่มีกรดอะมิโนเชื่อมต่อกันด้วยพันธะเปปไทด์ โครงสร้างหลักคือระดับที่ง่ายที่สุดของการจัดระเบียบโครงสร้างของโมเลกุลโปรตีน รับประกันความเสถียรสูงด้วยพันธะโควาเลนต์เปปไทด์ระหว่างกลุ่ม α-อะมิโนของกรดอะมิโนหนึ่งตัวกับกลุ่ม α-คาร์บอกซิลของกรดอะมิโนอีกตัวหนึ่ง
ในแง่ของพันธะเปปไทด์หากกลุ่มอะมิโนของโพรลีนหรือไฮดรอกซีโพรลีนเข้ามามีส่วนร่วมก็จะดูแตกต่างออกไป

เมื่อพันธะเปปไทด์เกิดขึ้นในไต กลุ่มคาร์บอกซิลของกรดอะมิโนหนึ่งตัวจะถูกกระตุ้น จากนั้นมันจะรวมตัวกับกลุ่มอะมิโนอีกกลุ่มหนึ่ง นอกจากนี้ยังสามารถทำการสังเคราะห์โพลีเปปไทด์ในห้องปฏิบัติการได้
ลิงค์เปปไทด์คือชิ้นส่วนของโพลีเปปไทด์แลนซ์ที่ทำซ้ำตัวเอง มีลักษณะเฉพาะบางประการที่เกี่ยวข้องไม่เพียงแต่กับรูปแบบของโครงสร้างหลักเท่านั้น แต่ยังรวมถึงระดับที่สำคัญที่สุดของการจัดระเบียบของมีดหมอโพลีเปปไทด์ด้วย:


· Coplanarity - อะตอมทั้งหมดที่รวมอยู่ในกลุ่มเปปไทด์นั้นอยู่ในระนาบเดียวกัน
· ความถูกต้องมีอยู่ในรูปแบบเรโซแนนซ์สองรูปแบบ (รูปแบบคีโตและอีนอล)
· ตำแหน่งทรานส์ของผู้วิงวอนในการเชื่อมต่อ C-N หนึ่งร้อยเปอร์เซ็นต์
· มีการก่อตัวของเอ็นน้ำ โดยที่ผิวหนังที่มีกลุ่มเปปไทด์สามารถสร้างเอ็นน้ำ 2 เส้นร่วมกับกลุ่มอื่นได้ รวมถึงกลุ่มเปปไทด์ด้วย
กลุ่มเปปไทด์เกี่ยวข้องกับกลุ่มอะมิโนของโพรลีนหรือไฮดรอกซีโพรลีน กลิ่นเหม็นของอาคารถูกสร้างขึ้นด้วยเสียงน้ำเพียงครั้งเดียว (สิ่งมหัศจรรย์) สิ่งนี้ระบุไว้บนโครงสร้างทุติยภูมิที่ขึ้นรูปของโปรตีน สารประกอบโพลีเปปไทด์ซึ่งประกอบด้วยโพรลีนหรือไฮดรอกซีโพรลีน ถูกทำลายได้ง่ายและไม่สามารถกำจัดออกได้ด้วยพันธะน้ำอื่นๆ เช่นเคย
รูปแบบการกำหนดสูตรไตรเปปไทด์:

ระดับของการจัดระเบียบเชิงพื้นที่ของโปรตีน: โครงสร้างรองของโปรตีน: แนวคิดของα-helix และ β-folded ball โครงสร้างระดับตติยภูมิของโปรตีน: แนวคิดเกี่ยวกับโปรตีนพื้นเมืองและการสูญเสียสภาพโปรตีน โครงสร้างควอเทอร์นารีของโปรตีนคล้ายกับฮีโมโกลบิน
โครงสร้างรองของโปรตีนภายใต้โครงสร้างทุติยภูมิของโปรตีน เราเข้าใจวิธีการจัดเรียงโพลีเปปไทด์แลนซ์ในโครงสร้างที่ได้รับคำสั่ง ตามการกำหนดค่าจะมองเห็นองค์ประกอบต่อไปนี้ของโครงสร้างรอง: α -เกลียว β - ลูกชิ้นอะไหล่โกดัง
โมเดลบูโดวี α-เกลียว ซึ่งปกป้องพลังทั้งหมดของพันธะเปปไทด์ ถูกทำลายโดย L. Pauling และ R. Corey (1949 - 1951)
ถึงลูกน้อย 3 กแผนภาพที่แสดง α -เกลียวซึ่งให้ข้อมูลเกี่ยวกับพารามิเตอร์หลักของมีดหมอโพลีเปปไทด์ที่ถูกเผาไหม้ α - เกลียวในลักษณะที่การหมุนของเกลียวสม่ำเสมอ ดังนั้นโครงเกลียวจึงมีความสมมาตรของสกรู (รูปที่ 3, ข). เมื่อถึงรอบหนัง α - เกลียวมีกรดอะมิโนส่วนเกิน 3.6 ยืนระหว่างทางเลี้ยวหรือขอบของเกลียวเป็น 0.54 นาโนเมตร โดยที่ทางเลี้ยวถัดไปถึง 26 ° การขึ้นรูปและการตัดแต่ง α - โครงร่างของเกลียวเกิดขึ้นจากโครงสร้างของเอ็นที่เป็นน้ำซึ่งถูกสร้างขึ้นระหว่างกลุ่มเปปไทด์ของผิวหนัง n-ไปซะ ( ป+3) กรดอะมิโนส่วนเกิน แม้ว่าพลังงานของเอ็นน้ำจะมีน้อย แต่มีปริมาณของเอ็นน้ำสูงจึงทำให้เกิดพลังอันสำคัญซึ่งเป็นผลมาจากการที่ α - โครงแบบเกลียวเพื่อทำให้ขาตั้งสมบูรณ์ อนุมูลทางชีวภาพของกรดอะมิโนส่วนเกินไม่ได้มีส่วนร่วมในสาร α - โครงสร้างแบบเฮลิคอล ดังนั้นกรดอะมิโนทั้งหมดจึงมีส่วนเกินเข้ามา α -เกลียวมีความเท่ากัน
โปรตีนจากธรรมชาตินั้นถนัดขวาน้อยกว่า α - เกลียว
ลูกบอลพับβ- องค์ประกอบอื่นของโครงสร้างรอง ในหน้าการบริหาร α -เกลียว β - ลูกบอลที่พับแล้วมีรูปทรงเป็นเส้นตรง ไม่ใช่รูปทรง Strizhnevuyu (รูปที่ 4) โครงสร้างเชิงเส้นถูกกำหนดโดยที่มาของพันธะน้ำระหว่างกลุ่มเปปไทด์ที่ยืนอยู่บนส่วนต่างๆ ของท่อโพลีเปปไทด์ แปลงเหล่านี้ดูเหมือนจะอยู่ใกล้กับพื้นผิวของจุดเชื่อมต่อน้ำระหว่าง - C = O และ HN - กลุ่ม (0.272 นาโนเมตร)

เล็ก 4. ภาพประกอบแผนผัง β
- โกดังบอลบ่อย (ลูกศรระบุ
 เกี่ยวกับมีดหมอโพลีเปปไทด์)
เกี่ยวกับมีดหมอโพลีเปปไทด์)
เล็ก 3. โครงการ ( ก) รุ่นนั้น ( ข) α -เกลียว
โครงสร้างรองของโปรตีนถูกระบุว่าเป็นโครงสร้างหลัก กรดอะมิโนส่วนเกินในปริมาณต่างๆ จะถูกสร้างขึ้นก่อนที่จะเกิดเอ็นน้ำแล้วจึงเทลงในส่วนผสม α -เกลียวหรือ β -ชารู กรดอะมิโนที่ละลายได้ในเกลียวประกอบด้วยอะลานีน กรดกลูตามิก กลูตามีน ลิวซีน ไลซีน เมไทโอนีน และฮิสทิดีน หากชิ้นส่วนโปรตีนถูกสร้างขึ้นจากกรดอะมิโนส่วนเกินเป็นหลัก ก็จะก่อตัวในขั้นตอนนี้ α -เกลียว. วาลีน ไอโซลิวซีน ธรีโอนีน ไทโรซีน และฟีนิลอะลานีนเป็นส่วนผสม β - ลูกบอลโพลีเปปไทด์มีดหมอ โครงสร้างที่ไม่เป็นระเบียบปรากฏบนแปลงของโพลีเปปไทด์แลนซ์ ซึ่งมีกรดอะมิโนตกค้าง เช่น ไกลซีน, ซีรีน, กรดแอสปาร์ติก, แอสพาราจีน, โพรลีนเข้มข้น
อุดมไปด้วยโปรตีนทันที α -เกลียวนั่น β -ชาริ ส่วนหนึ่งของโครงสร้างขดนั้นแตกต่างกันไปตามโปรตีนต่างๆ ดังนั้นพาราไมโอซินโปรตีนจากเนื้อสัตว์จึงถูกทำให้เป็นเกลียวเกือบ 100% สัดส่วนที่สูงของการกำหนดค่าขดลวดในไมโอโกลบินและฮีโมโกลบิน (75%) อย่างไรก็ตาม ในทริปซินและไรโบนิวคลีเอส ส่วนสำคัญของโพลีเปปไทด์ lanjug จะพอดีกับชารูเวต β -โครงสร้าง โปรตีนของเนื้อเยื่อรองรับ - เคราติน (โปรตีนของเส้นผม), คอลลาเจน (โปรตีนของผิวหนังและเอ็น) - บวม β - การกำหนดค่าของหอกโพลีเปปไทด์
โครงสร้างโปรตีนระดับตติยภูมิโครงสร้างระดับตติยภูมิของโปรตีนเป็นวิธีการวางหอกโพลีเปปไทด์ในอวกาศ เพื่อให้โปรตีนเพิ่มพลังให้กับพลังการทำงานของมัน มีดหมอโพลีเปปไทด์จำเป็นต้องเผาในอวกาศเพียงลำพัง ทำให้เกิดโครงสร้างที่ทำงานได้ตามหน้าที่ โครงสร้างนี้เรียกว่า พื้นเมือง. โดยไม่คำนึงถึงโครงสร้างเชิงพื้นที่จำนวนมากที่เป็นไปได้ในทางทฤษฎีสำหรับมีดหมอโพลีเปปไทด์ที่อยู่รอบๆ ควรนำโปรตีนกล่องเสียงมาสร้างโครงสร้างดั้งเดิมเดียว
ปรับโครงสร้างตติยภูมิของโปรตีนให้คงที่โดยปฏิกิริยาที่เกิดขึ้นระหว่างอนุมูลทางชีวภาพของกรดอะมิโนที่มากเกินไปในส่วนต่างๆ ของโพลีเปปไทด์แลนซ์ ปฏิสัมพันธ์เหล่านี้สามารถแบ่งออกเป็นจุดแข็งและจุดอ่อน
อันตรกิริยาที่รุนแรงเกี่ยวข้องกับพันธะโควาเลนต์ระหว่างอะตอมซัลเฟอร์ของซิสเทอีนส่วนเกิน ซึ่งอยู่ที่ส่วนต่างๆ ของแลนซ์โพลีเปปไทด์ มิฉะนั้น พันธะดังกล่าวจะเรียกว่าสะพานไดซัลไฟด์ การก่อตัวของสะพานซัลไฟด์สามารถแสดงได้ดังนี้:
นอกจากพันธะโควาเลนต์แล้ว โครงสร้างตติยภูมิของโมเลกุลโปรตีนยังได้รับอิทธิพลจากปฏิกิริยาที่อ่อนแอ ซึ่งโดยธรรมชาติแล้ว จะถูกแบ่งออกเป็นขั้วและไม่มีขั้ว
ปฏิกิริยาเชิงขั้วเกี่ยวข้องกับการเชื่อมต่อของไอออนและน้ำ อันตรกิริยาของไอออนิกถูกสร้างขึ้นจากการสัมผัสระหว่างกลุ่มของเสียที่มีประจุบวก ได้แก่ ไลซีน อาร์จินีน ฮิสทิดีน และกลุ่ม COOH ที่มีประจุลบของกรดแอสปาร์ติกและกลูตามิก การเชื่อมต่อของน้ำเกิดขึ้นระหว่างกลุ่มการทำงานของอนุมูลทางชีวภาพของกรดอะมิโนที่มากเกินไป
ปฏิกิริยาแบบไม่มีขั้วหรือแบบแวนเดอร์วาลส์ระหว่างอนุมูลคาร์โบไฮเดรตของกรดอะมิโนที่ตกค้างช่วยป้องกันการก่อตัว แกนที่ไม่ชอบน้ำ (จุดไขมัน) ที่อยู่ตรงกลางของโปรตีนโกลบูลเพราะว่า อนุมูลคาร์โบไฮเดรตจะหายไปเมื่อสัมผัสกับน้ำ ยิ่งโปรตีนมีกรดอะมิโนไม่มีขั้วมากเท่าใด บทบาทของพันธะแวนเดอร์วาลส์ในโครงสร้างตติยภูมิที่มีรูปร่างก็จะยิ่งมากขึ้นเท่านั้น
การเชื่อมต่อเชิงตัวเลขระหว่างอนุมูลโปรตีนของกรดอะมิโนส่วนเกินบ่งบอกถึงการกำหนดค่าเชิงพื้นที่ของโมเลกุลโปรตีน (รูปที่ 5)

เล็ก 5. ประเภทของเอ็นที่รองรับโครงสร้างตติยภูมิของโปรตีน:
ก- ส่วนผสมซัลไฟด์ ข -ลิงค์อิออน; ซีดี -การเชื่อมต่อน้ำ
ง -พันธบัตรของฟาน เดอร์ วาลส์
โครงสร้างระดับตติยภูมิของโปรตีนที่ได้รับมาใหม่มีลักษณะเฉพาะ เช่นเดียวกับโครงสร้างหลักที่มีเอกลักษณ์เฉพาะตัว เฉพาะพื้นที่ที่ถูกต้องในการวางโปรตีนเท่านั้นที่จะทำให้โปรตีนยังคงทำงานอยู่ การทำลายโครงสร้างที่สามครั้งใหญ่ทำให้เกิดการเปลี่ยนแปลงระดับโปรตีนและการสูญเสียกิจกรรมทางชีวภาพ
โครงสร้างควอเทอร์นารีของโปรตีนโปรตีนที่มีน้ำหนักโมเลกุลมากกว่า 100 kDa 1 มักจะประกอบด้วยโพลีเปปไทด์หลายอันที่มีน้ำหนักโมเลกุลน้อย โครงสร้างซึ่งประกอบด้วยหอกโพลีเปปไทด์จำนวนมากซึ่งครอบครองตำแหน่งคงที่อย่างเคร่งครัดซึ่งเห็นได้ชัดว่าเป็นอีกตำแหน่งหนึ่งซึ่งเป็นผลมาจากการที่โปรตีนมีกิจกรรมที่แตกต่างกันเรียกว่าโครงสร้างควอเทอร์นารีของโปรตีน โปรตีนที่มีโครงสร้างหนึ่งในสี่เรียกว่า โมเลกุลหรืออย่างอื่น มัลติเมอร์ และโกดังของมีดหมอโพลีเปปไทด์นั้นชัดเจน หน่วยย่อย หรืออย่างอื่น โปรโตเมียร์ . พลังลักษณะเฉพาะของโปรตีนที่มีโครงสร้างควอเทอร์นารีคือโปรตีนที่หน่วยย่อยไม่มีฤทธิ์ทางชีวภาพ
การรักษาเสถียรภาพของโครงสร้างควอเทอร์นารีของโปรตีนนั้นเกิดขึ้นได้จากปฏิกิริยาของปฏิกิริยาเชิงขั้วระหว่างอนุมูลของกรดอะมิโนที่อยู่บนพื้นผิวของหน่วยย่อย การโต้ตอบดังกล่าวมีความสำคัญในการลดหน่วยย่อยของคอมเพล็กซ์ที่จัดไว้ การแบ่งหน่วยย่อยที่มีปฏิสัมพันธ์กันเรียกว่าผู้ติดต่อสาวใช้
ตัวอย่างคลาสสิกของโปรตีนที่มีโครงสร้างหนึ่งในสี่คือฮีโมโกลบิน โมเลกุลของฮีโมโกลบินมีน้ำหนักโมเลกุล 68,000 ประกอบด้วยหน่วยย่อย 4 หน่วยจาก 2 ประเภทที่แตกต่างกัน α і β / α -หน่วยย่อยประกอบด้วยกรดอะมิโน 141 ชนิด ก β - iz 146 โครงสร้างระดับตติยภูมิ α - І β -หน่วยย่อยคล้ายกับน้ำหนักโมเลกุล (17,000 ดังนั้น) หน่วยย่อยผิวหนังแก้แค้นกลุ่มขาเทียม - ฮีม . ชิ้นส่วนของฮีมยังอยู่ในโปรตีนอื่น ๆ (ไซโตโครม, ไมโอโกลบิน) ซึ่งก่อตัวเพิ่มเติม แต่เราอยากจะพูดคุยสั้น ๆ ถึงโครงสร้างของโปรตีนเหล่านั้น (รูปที่ 6) ฮีมที่จัดกลุ่มเป็นระบบวงจรโคพลานาร์แบบพับที่ประกอบด้วยอะตอมกลาง ซึ่งสร้างพันธะประสานงานกับโพลีเมอร์ส่วนเกินบางส่วน เชื่อมต่อกันด้วยตำแหน่งมีเทน (= CH -) ในเฮโมโกลบินระดับออกซิเดชันเริ่มเกิดขึ้น (2+)
หลายหน่วยย่อย - สอง α และสอง β - รวมเป็นโครงสร้างเดียวในลักษณะที่ α -หน่วยย่อยติดต่อเท่านั้น β -หน่วยย่อยและโดยรวม (รูปที่ 7)

เล็ก 6. โครงสร้างของฮีโมโกลบิน

เล็ก 7. แผนผังแสดงโครงสร้างควอเทอร์นารีของฮีโมโกลบิน:
เฟ - ฮีมเฮโมโกลบิน
ดังที่เห็นได้จากเด็กทารกวัย 7 ขวบ ฮีโมโกลบิน 1 โมเลกุลสามารถขนส่งกรดได้ 4 โมเลกุล ทั้งการจับและการปลดปล่อยความเป็นกรดจะมาพร้อมกับการเปลี่ยนแปลงโครงสร้างในโครงสร้าง α - І β -หน่วยย่อยของฮีโมโกลบินและการสลายตัวร่วมกันในอีพิโมเลกุล ข้อเท็จจริงนี้ยืนยันว่าโครงสร้างไตรมาสของโปรตีนไม่เข้มงวดอีกต่อไป
ข้อมูลที่คล้ายกัน.
คุณสมบัติอย่างหนึ่งของโปรตีนคือการจัดโครงสร้างที่ซับซ้อน โปรตีนทั้งหมดมีโครงสร้างปฐมภูมิ ทุติยภูมิ และตติยภูมิ และโปรตีนที่มี PPC สองตัวขึ้นไปจะมีโครงสร้างควอเทอร์นารี (QS)
โครงสร้างโปรตีนปฐมภูมิ (PSB) – นี่คือลำดับของการสะสม (ลำดับ) ของกรดอะมิโนส่วนเกินใน PPC.
อย่างไรก็ตาม เนื่องจากความอุดมสมบูรณ์และองค์ประกอบของกรดอะมิโน โปรตีนจึงมีหลายประเภท ตัวอย่างเช่น จากกรดอะมิโน 2 ตัว คุณสามารถรวมไดเปปไทด์ 2 ชนิดเข้าด้วยกันได้:
เมื่อจำนวนกรดอะมิโนมากกว่า 20 จำนวนการผสมที่เป็นไปได้จะน้อยกว่า 210 18 . และถ้าคุณรู้ว่าใน PPC กรดอะมิโนของผิวหนังสามารถไวต่อความรู้สึกได้มากกว่าหนึ่งครั้ง การปกป้องตัวเลือกที่เป็นไปได้มากมายจึงเป็นเรื่องสำคัญ
ความสำคัญของโครงสร้างโปรตีนปฐมภูมิ (PPS)
สามารถคำนวณ PSB ของโปรตีนเพื่อขอความช่วยเหลือได้ ฟีนิลไทโอไฮแดนโทอิน
วิธี
. วิธีนี้ขึ้นอยู่กับปฏิกิริยาระหว่างกัน ฟีนิลไอโซไทโอไซยาเนต
(FITC) ด้วย α-AA เป็นผลให้เกิดความสัมพันธ์ที่ซับซ้อนขึ้น FITC-AK
.
ตัวอย่างเช่น ลองดูที่เปปไทด์ ![]() เมื่อใช้วิธีการนี้ PSB จะถูกนำมาใช้เพื่อกำหนดลำดับของกรดอะมิโนส่วนเกิน
เมื่อใช้วิธีการนี้ PSB จะถูกนำมาใช้เพื่อกำหนดลำดับของกรดอะมิโนส่วนเกิน
FITC ทำปฏิกิริยากับกรดอะมิโนส่วนปลาย (a) กำลังจัดตั้งคอมเพล็กซ์ FTG-กซึ่งถูกเติมลงในส่วนผสมและบ่งบอกถึงประสิทธิภาพของกรดอะมิโน ก. ตัวอย่างเช่น tse - asn
![]() ฯลฯ เสริมสร้างและระบุกรดอะมิโนอื่นๆ ทั้งหมดอย่างสม่ำเสมอ มันเป็นกระบวนการที่ลำบาก โปรตีน PSB ขนาดกลางคงอยู่ได้นานหลายเดือน
ฯลฯ เสริมสร้างและระบุกรดอะมิโนอื่นๆ ทั้งหมดอย่างสม่ำเสมอ มันเป็นกระบวนการที่ลำบาก โปรตีน PSB ขนาดกลางคงอยู่ได้นานหลายเดือน
การถอดรหัส PSB มีความสำคัญ เซนเกรุ(1953) ซึ่งเป็นการค้นพบอินซูลิน PSB (ผู้ได้รับรางวัลโนเบล) โมเลกุลอินซูลินประกอบด้วย 2 PPCs - A และ B

A-lancet ประกอบด้วยกรดอะมิโน 21 ตัว มีดหมอ - จาก 30 ตัว PPC เชื่อมต่อกันโดยไซต์ซัลไฟด์ จำนวนโปรตีนที่มีการระบุ PSB ในวันนี้ถึง 1,500 การเปลี่ยนแปลงเล็กน้อยในโครงสร้างหลักสามารถเปลี่ยนพลังของโปรตีนได้อย่างมาก ในเม็ดเลือดแดงของคนที่มีสุขภาพดี HbA มีอยู่ - เมื่อแทนที่ใน -lancer HbA ที่ตำแหน่งที่ 6 กาวบน เพลาโทษของการเจ็บป่วยร้ายแรง โรคโลหิตจางเซลล์เคียวสำหรับเด็กคนใดที่เกิดมาพร้อมกับความผิดปกตินี้ให้เสียชีวิตก่อนกำหนด ในทางกลับกัน มีตัวเลือกที่เป็นไปได้ในการเปลี่ยน PSB ซึ่งไม่ได้ระบุไว้โดยหน่วยงานด้านเคมีกายภาพและชีวภาพ ตัวอย่างเช่น, HbC อยู่ในตำแหน่งที่ 6 ของ b-lance แทนที่จะเป็น glu - liz, HbC อาจไม่แข่งขันกับอำนาจเหนือ HbA และผู้ที่มี Hb ดังกล่าวในเม็ดเลือดแดงจะมีสุขภาพที่ดีในทางปฏิบัติ
ความเสถียรของ PSBมีหลักประกันโดยพันธะโควาเลนต์เปปไทด์ภายใน และอีกนัยหนึ่งคือพันธะไดซัลไฟด์
โครงสร้างทุติยภูมิของโปรตีน (PSB)
PPV ของโปรตีนแสดงความยืดหยุ่นอย่างมากและพัฒนาโครงสร้างที่กว้างขวางหรือโดดเด่น โครงสร้าง. ในโปรตีน โครงสร้างดังกล่าวมี 2 ระดับ คือ VSB และโครงสร้างตติยภูมิ (แจ้งภายหลัง)
วีเอสบี – การกำหนดค่าของ PPC นี้ วิธีการวางหรือบิดเป็นโครงสร้างใด ๆ สอดคล้องกับโปรแกรมที่มีอยู่ใน PPCเอสบี.
VSB มีสามประเภทหลัก:
1) -เกลียว;
2) ข-โครงสร้าง(สต็อกชิ้นส่วนลูกหรือแผ่นชิ้นส่วนพับ);
3) ลูกบอลไร้กังวล
-เกลียว .
แบบจำลองนี้เสนอโดย V. Pauling วิธีนี้เหมาะที่สุดสำหรับโปรตีนทรงกลม สำหรับระบบใดๆ สถานะที่เสถียรที่สุดคือสถานะที่แสดงถึงพลังงานอิสระขั้นต่ำ สำหรับเปปไทด์ สถานการณ์นี้เกิดขึ้นเมื่อกลุ่ม CO และ NH เชื่อมต่อกันด้วยพันธะน้ำที่อ่อนแอ ยู ก -เกลียว กลุ่ม NH ของกรดอะมิโนตัวที่ 1 ตกค้างมีปฏิกิริยากับกลุ่ม CO ของกรดอะมิโนตัวที่ 4 เป็นผลให้แกนหลักของเปปไทด์สร้างเกลียวซึ่งมีส่วนเกิน 3.6 AA ตกอยู่ที่การหมุนของผิวหนัง
เกลียวเกลียว 1 เส้น (1 รอบ) = 3.6 AC = 0.54 นาโนเมตร ตัดเป็นเกลียว – 26°
การบิดของ PPC จะสังเกตได้ด้านหลังลูกศรปี เพื่อให้เกลียวเคลื่อนไปทางขวา ผ่านผิวหนัง 5 รอบ (18 AC; 2.7 นาโนเมตร) การกำหนดค่า PPC จะถูกทำซ้ำ
มีเสถียรภาพ วีเอสบีประการแรกก่อนพันธะน้ำและอีกประการหนึ่ง - เปปไทด์และซัลไฟด์ พันธะน้ำอ่อนกว่าพันธะเคมีมาตรฐานถึง 10-100 เท่า อย่างไรก็ตาม กลิ่นเหม็นในปริมาณมากจะช่วยรับประกันความกระด้างและความแน่นของ VSB R-lancelets มีลักษณะเป็นเกลียว เอียงจนถึงจุดที่มีเสียงดัง และกระจายไปตามด้านและแกนต่างๆ
ข -โครงสร้าง .
เหล่านี้เป็นส่วนที่พับของโครง PPC ซึ่งมีลักษณะคล้ายใบไม้พับเหมือนหีบเพลง ลูกบอลของ PPC สามารถขนานกันได้เนื่องจากการรุกเริ่มต้นที่ N- หรือ C-end
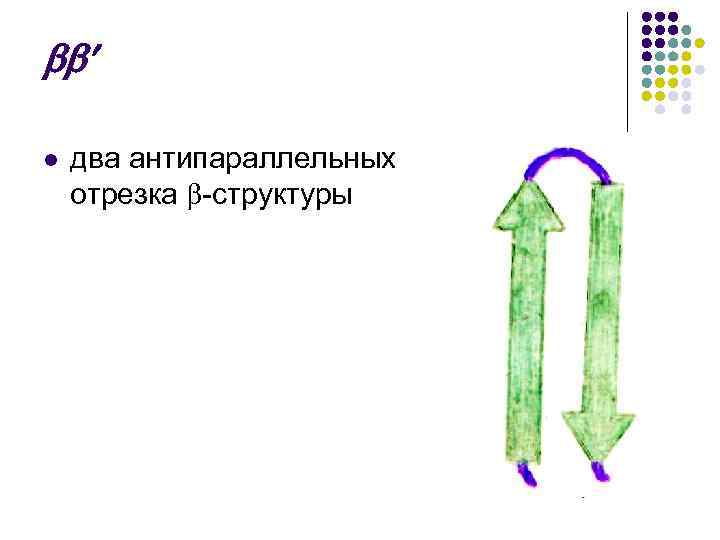
เนื่องจากหอกแคบของลูกบอลมีทิศทางที่ปลายใกล้เคียง N–C และ C–N ดังนั้นจึงเรียกว่ากลิ่นเหม็น ต่อต้านขนาน.

 ขนาน
ขนาน
![]() ต่อต้านขนาน
ต่อต้านขนาน
การก่อตัวของพันธะน้ำเกิดขึ้นเช่นเดียวกับใน a-helix ระหว่างกลุ่ม CO และ NH

 การศึกษาการจัดโครงสร้างของโปรตีนเป็นหนึ่งในปัญหาหลักของชีวเคมีสมัยใหม่ มีความสำคัญทางวิทยาศาสตร์และการปฏิบัติที่สำคัญสำหรับการทำความเข้าใจการขยายตัวอย่างมากของการทำงานของโปรตีน มีกรดอะมิโนหลายสิบหรือหลายร้อยตัวในโมเลกุลโปรตีนหรือไม่?
การศึกษาการจัดโครงสร้างของโปรตีนเป็นหนึ่งในปัญหาหลักของชีวเคมีสมัยใหม่ มีความสำคัญทางวิทยาศาสตร์และการปฏิบัติที่สำคัญสำหรับการทำความเข้าใจการขยายตัวอย่างมากของการทำงานของโปรตีน มีกรดอะมิโนหลายสิบหรือหลายร้อยตัวในโมเลกุลโปรตีนหรือไม่?
 โปรตีนเอมิลฟิชเชอร์เป็นโพลีเปปไทด์แบบพับซึ่งมีกรดอะมิโนเกาะกันด้วยพันธะเปปไทด์ (R-CO-NH-R) ซึ่งก่อตัวขึ้นระหว่างปฏิกิริยาระหว่างคาร์บอกซิลและกลุ่มอะมิโนของกรดอะมิโน
โปรตีนเอมิลฟิชเชอร์เป็นโพลีเปปไทด์แบบพับซึ่งมีกรดอะมิโนเกาะกันด้วยพันธะเปปไทด์ (R-CO-NH-R) ซึ่งก่อตัวขึ้นระหว่างปฏิกิริยาระหว่างคาร์บอกซิลและกลุ่มอะมิโนของกรดอะมิโน
 ข้อพิสูจน์เชิงทดลองของทฤษฎี l l l l โปรตีนธรรมชาติมีหมู่ไตเตรทจำนวนน้อย – COOH และ –NH 2 ในระหว่างกระบวนการไฮโดรไลซิสของโปรตีน จำนวนกลุ่มไตเตรท – COOH และ –NH 2 ถูกสร้างขึ้นภายใต้การกระทำของโปรตีโอไลต์ในจำนวนปริมาณสัมพันธ์ ใน การปรากฏตัวของเอนไซม์บางชนิดโปรตีนจะถูกแบ่งออกเป็นชิ้นส่วนที่กำหนดอย่างเคร่งครัด (โพลีเปปไทด์) การวิเคราะห์ด้วยรังสีเอกซ์ยืนยันโครงสร้างโพลีเปปไทด์ โปรตีน การยืนยันหลักของทฤษฎีโพลีเปปไทด์ของโปรตีนคือความเป็นไปได้ในการสังเคราะห์โพลีเปปไทด์และโปรตีนจากที่รู้จักอยู่แล้ว โปรตีน (อินซูลิน, ไลโซไซม์, ไรโบนิวเคลียส) โดยวิธีทางเคมี
ข้อพิสูจน์เชิงทดลองของทฤษฎี l l l l โปรตีนธรรมชาติมีหมู่ไตเตรทจำนวนน้อย – COOH และ –NH 2 ในระหว่างกระบวนการไฮโดรไลซิสของโปรตีน จำนวนกลุ่มไตเตรท – COOH และ –NH 2 ถูกสร้างขึ้นภายใต้การกระทำของโปรตีโอไลต์ในจำนวนปริมาณสัมพันธ์ ใน การปรากฏตัวของเอนไซม์บางชนิดโปรตีนจะถูกแบ่งออกเป็นชิ้นส่วนที่กำหนดอย่างเคร่งครัด (โพลีเปปไทด์) การวิเคราะห์ด้วยรังสีเอกซ์ยืนยันโครงสร้างโพลีเปปไทด์ โปรตีน การยืนยันหลักของทฤษฎีโพลีเปปไทด์ของโปรตีนคือความเป็นไปได้ในการสังเคราะห์โพลีเปปไทด์และโปรตีนจากที่รู้จักอยู่แล้ว โปรตีน (อินซูลิน, ไลโซไซม์, ไรโบนิวเคลียส) โดยวิธีทางเคมี

 ลักษณะโครงสร้างของเปปไทด์แลนซ์ l l การเชื่อมโยงเปปไทด์สั้นกว่าการเชื่อมโยง –C–N– ประมาณ 10% และมีลักษณะเป็นการเชื่อมโยง “ที่จมอยู่ใต้น้ำบ่อยครั้ง” –C=N– โดย L. Pauling และ R. Corey ซึ่ง พัฒนาในปี 1948–1955 หน้า อธิบายลักษณะพิเศษของพันธะ C-N ด้วย "การสั่นพ้อง" ระหว่างทั้งสองรูปแบบ
ลักษณะโครงสร้างของเปปไทด์แลนซ์ l l การเชื่อมโยงเปปไทด์สั้นกว่าการเชื่อมโยง –C–N– ประมาณ 10% และมีลักษณะเป็นการเชื่อมโยง “ที่จมอยู่ใต้น้ำบ่อยครั้ง” –C=N– โดย L. Pauling และ R. Corey ซึ่ง พัฒนาในปี 1948–1955 หน้า อธิบายลักษณะพิเศษของพันธะ C-N ด้วย "การสั่นพ้อง" ระหว่างทั้งสองรูปแบบ
 ลักษณะโครงสร้างของเปปไทด์แลนซ์ l l l l กล่าวอีกนัยหนึ่ง ในโปรตีนและเปปไทด์ พันธะ C-N มักจะทวีคูณผ่านอันตรกิริยาของอิเล็กตรอนคู่เดียวของอะตอมไนโตรเจนกับระบบอิเล็กทรอนิกส์ของหมู่คาร์บอนิล ซึ่งนำไปสู่ความยากลำบาก การก่อตัว tannya navkolo zvjazku C–N Zazvichay peptide zv' รูปแบบทรานส์จะเด่นชัดกว่ารูปแบบซิสประมาณ 2.6 kcal/mol (10.878 kJ/mol) เนื่องจากความใกล้ชิดของอะตอมคาร์บอนในรูปแบบซิสทำให้ยากต่อการ
ลักษณะโครงสร้างของเปปไทด์แลนซ์ l l l l กล่าวอีกนัยหนึ่ง ในโปรตีนและเปปไทด์ พันธะ C-N มักจะทวีคูณผ่านอันตรกิริยาของอิเล็กตรอนคู่เดียวของอะตอมไนโตรเจนกับระบบอิเล็กทรอนิกส์ของหมู่คาร์บอนิล ซึ่งนำไปสู่ความยากลำบาก การก่อตัว tannya navkolo zvjazku C–N Zazvichay peptide zv' รูปแบบทรานส์จะเด่นชัดกว่ารูปแบบซิสประมาณ 2.6 kcal/mol (10.878 kJ/mol) เนื่องจากความใกล้ชิดของอะตอมคาร์บอนในรูปแบบซิสทำให้ยากต่อการ
 ลักษณะโครงสร้างของเปปไทด์แลนซ์ l l l l สามารถหมุนได้รอบพันธะอย่างง่าย 2 พันธะ (N–C และ C–C 1) ซึ่งอยู่ติดกับอะตอมที่ไม่สมมาตร = 180 - ทรานส์ฟอร์ม)
ลักษณะโครงสร้างของเปปไทด์แลนซ์ l l l l สามารถหมุนได้รอบพันธะอย่างง่าย 2 พันธะ (N–C และ C–C 1) ซึ่งอยู่ติดกับอะตอมที่ไม่สมมาตร = 180 - ทรานส์ฟอร์ม)
 คุณสมบัติโครงสร้างของมีดหมอเปปไทด์ l l เนื่องจากปฏิสัมพันธ์ระหว่างผู้พิทักษ์ในมีดหมอเปปไทด์ พวกเขาไม่สามารถมีความหมายใด ๆ ได้ - อนุญาตให้มีเพียงไม่กี่พื้นที่ที่แยกจากกันเท่านั้นซึ่งบ่งบอกถึงความแข็งแกร่งที่มีพลัง ความสอดคล้องบางประการของเปปไทด์ lanjug Kuti มีปฏิสัมพันธ์กับแต่ละอัน อื่น ๆ การเปลี่ยนแปลงหนึ่งในนั้นยากกว่าการเปลี่ยนแปลงอีก
คุณสมบัติโครงสร้างของมีดหมอเปปไทด์ l l เนื่องจากปฏิสัมพันธ์ระหว่างผู้พิทักษ์ในมีดหมอเปปไทด์ พวกเขาไม่สามารถมีความหมายใด ๆ ได้ - อนุญาตให้มีเพียงไม่กี่พื้นที่ที่แยกจากกันเท่านั้นซึ่งบ่งบอกถึงความแข็งแกร่งที่มีพลัง ความสอดคล้องบางประการของเปปไทด์ lanjug Kuti มีปฏิสัมพันธ์กับแต่ละอัน อื่น ๆ การเปลี่ยนแปลงหนึ่งในนั้นยากกว่าการเปลี่ยนแปลงอีก

 l K. W. Linderström-Lang แยกแยะการจัดระเบียบโมเลกุลโปรตีนได้ 4 ระดับ - โครงสร้างหลัก, ทุติยภูมิ, ตติยภูมิและควอเทอร์นารี แม้ว่าหมวดหมู่เหล่านี้จะล้าสมัยแล้ว แต่จะยังคงใช้ต่อไปในขณะนี้
l K. W. Linderström-Lang แยกแยะการจัดระเบียบโมเลกุลโปรตีนได้ 4 ระดับ - โครงสร้างหลัก, ทุติยภูมิ, ตติยภูมิและควอเทอร์นารี แม้ว่าหมวดหมู่เหล่านี้จะล้าสมัยแล้ว แต่จะยังคงใช้ต่อไปในขณะนี้

 โครงสร้างหลักของโปรตีน ลำดับของกรดอะมิโนที่ตกค้างในมีดหมอโพลีเปปไทด์ถูกเข้ารหัสโดยยีนโครงสร้างของโปรตีนนี้ และมีทุกสิ่งที่จำเป็นสำหรับการจัดระเบียบตนเองของโครงสร้างที่กว้างขวาง โปรตีนทั้งหมดแข่งขันกันเพื่อตัวมันเอง ด้วยโครงสร้างหลักนี้ จำนวนศักยภาพของโครงสร้างดังกล่าวไม่ได้จำกัดอยู่ที่จำนวนโปรตีนชนิดต่าง ๆ ในสิ่งมีชีวิตทุกประเภท ประมาณปี 1010-1012
โครงสร้างหลักของโปรตีน ลำดับของกรดอะมิโนที่ตกค้างในมีดหมอโพลีเปปไทด์ถูกเข้ารหัสโดยยีนโครงสร้างของโปรตีนนี้ และมีทุกสิ่งที่จำเป็นสำหรับการจัดระเบียบตนเองของโครงสร้างที่กว้างขวาง โปรตีนทั้งหมดแข่งขันกันเพื่อตัวมันเอง ด้วยโครงสร้างหลักนี้ จำนวนศักยภาพของโครงสร้างดังกล่าวไม่ได้จำกัดอยู่ที่จำนวนโปรตีนชนิดต่าง ๆ ในสิ่งมีชีวิตทุกประเภท ประมาณปี 1010-1012
 l l l l เป็นไปไม่ได้ที่จะเข้าใจการทำงานทางชีววิทยาและกลไกระดับโมเลกุลของกิจกรรมทางสรีรวิทยาของโปรตีนโดยปราศจากความรู้โดยละเอียดเกี่ยวกับการมีอยู่ของมัน การตรวจสอบโครงสร้างหลักของโปรตีนที่ "ก่อกลายพันธุ์" ทำให้สามารถเข้าใจธรรมชาติของโรคภาวะถดถอยในระดับโมเลกุลได้
l l l l เป็นไปไม่ได้ที่จะเข้าใจการทำงานทางชีววิทยาและกลไกระดับโมเลกุลของกิจกรรมทางสรีรวิทยาของโปรตีนโดยปราศจากความรู้โดยละเอียดเกี่ยวกับการมีอยู่ของมัน การตรวจสอบโครงสร้างหลักของโปรตีนที่ "ก่อกลายพันธุ์" ทำให้สามารถเข้าใจธรรมชาติของโรคภาวะถดถอยในระดับโมเลกุลได้
 วิธีการกำหนดโครงสร้างหลัก l l การเตรียมโปรตีนก่อนการวิเคราะห์โครงสร้างหลักมีวัตถุประสงค์เพื่อลดการซึมซาบของโครงสร้างอื่น ๆ ที่สูงกว่า กล่าวอีกนัยหนึ่ง หัวข้อของการวิเคราะห์อาจทำให้แลนซ์โปรตีนนี้ไม่เป็นระเบียบโดยไม่มี พันธะโควาเลนต์ตามขวาง (เช่น ไดซัลไฟด์) เพื่อให้พันธะเปปไทด์ทั้งหมดสามารถใช้ได้กับทั้งตัวทำปฏิกิริยาเคมีและเอนไซม์
วิธีการกำหนดโครงสร้างหลัก l l การเตรียมโปรตีนก่อนการวิเคราะห์โครงสร้างหลักมีวัตถุประสงค์เพื่อลดการซึมซาบของโครงสร้างอื่น ๆ ที่สูงกว่า กล่าวอีกนัยหนึ่ง หัวข้อของการวิเคราะห์อาจทำให้แลนซ์โปรตีนนี้ไม่เป็นระเบียบโดยไม่มี พันธะโควาเลนต์ตามขวาง (เช่น ไดซัลไฟด์) เพื่อให้พันธะเปปไทด์ทั้งหมดสามารถใช้ได้กับทั้งตัวทำปฏิกิริยาเคมีและเอนไซม์
 l l โปรตีนมีหน้าที่รับผิดชอบในการสูญเสียสภาพธรรมชาติเชิงลึกทั้งหมดและการสูญเสียควอเทอร์นารี ตติยภูมิ และหากเป็นไปได้ โครงสร้างทุติยภูมิ
l l โปรตีนมีหน้าที่รับผิดชอบในการสูญเสียสภาพธรรมชาติเชิงลึกทั้งหมดและการสูญเสียควอเทอร์นารี ตติยภูมิ และหากเป็นไปได้ โครงสร้างทุติยภูมิ
 l เพื่อป้องกันการเกิดพันธะไดซัลไฟด์ ให้ปิดกั้นหมู่ซัลไฮดริลอีกครั้งด้วยกรดไอโอโดกติกส่วนเกิน: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI
l เพื่อป้องกันการเกิดพันธะไดซัลไฟด์ ให้ปิดกั้นหมู่ซัลไฮดริลอีกครั้งด้วยกรดไอโอโดกติกส่วนเกิน: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI
 จากนั้นจะทำการคัดเลือกไฮโดรไลซิสของโปรตีนที่ถูกทำให้เสียสภาพเพื่อกำจัดระบบเปปไทด์ที่ทับซ้อนกัน การไฮโดรไลซิสดำเนินการด้วยเอนไซม์: l l l l trypsin (การจับที่สร้างโดยกลุ่มคาร์บอกซิลของกรดอะมิโนอะโรมาติก), chymotrypsin (การจับที่สร้างโดยกลุ่ม -COOH ของกรดอะมิโนอะโรมาติก) และอื่น ๆ ; รีเอเจนต์สารเคมี: โบรโมไซยาน์ Br - C N (สารยึดเกาะที่ประกอบขึ้นด้วยหมู่ - พบ COOH) เป็นต้น
จากนั้นจะทำการคัดเลือกไฮโดรไลซิสของโปรตีนที่ถูกทำให้เสียสภาพเพื่อกำจัดระบบเปปไทด์ที่ทับซ้อนกัน การไฮโดรไลซิสดำเนินการด้วยเอนไซม์: l l l l trypsin (การจับที่สร้างโดยกลุ่มคาร์บอกซิลของกรดอะมิโนอะโรมาติก), chymotrypsin (การจับที่สร้างโดยกลุ่ม -COOH ของกรดอะมิโนอะโรมาติก) และอื่น ๆ ; รีเอเจนต์สารเคมี: โบรโมไซยาน์ Br - C N (สารยึดเกาะที่ประกอบขึ้นด้วยหมู่ - พบ COOH) เป็นต้น
 l l l ขั้นตอนแรก - การแยกส่วนของเปปไทด์ที่แยกได้ - ดำเนินการโดยวิธีโครมาโตกราฟี ขั้นตอนที่เหลือ - การกำหนดโครงสร้างหลัก l เริ่มต้นด้วยความสำคัญของน้ำหนักโมเลกุลองค์ประกอบของกรดอะมิโน N- และกรดอะมิโนส่วนเกินที่ปลาย และคัดเลือกอีกชนิดที่แตกต่างจากชิ้นส่วนเปปไทด์ชุดแรกมาแบ่งและวิเคราะห์เพื่อกำหนดลำดับการกระจายตัวของกรดอะมิโนในโปรตีน
l l l ขั้นตอนแรก - การแยกส่วนของเปปไทด์ที่แยกได้ - ดำเนินการโดยวิธีโครมาโตกราฟี ขั้นตอนที่เหลือ - การกำหนดโครงสร้างหลัก l เริ่มต้นด้วยความสำคัญของน้ำหนักโมเลกุลองค์ประกอบของกรดอะมิโน N- และกรดอะมิโนส่วนเกินที่ปลาย และคัดเลือกอีกชนิดที่แตกต่างจากชิ้นส่วนเปปไทด์ชุดแรกมาแบ่งและวิเคราะห์เพื่อกำหนดลำดับการกระจายตัวของกรดอะมิโนในโปรตีน

 วิธีการของ F. Sanger l U 1945 r. นักชีวเคมีชาวอังกฤษ F. Sanger ได้พัฒนาวิธีแรกในการระบุกรดอะมิโนที่ปลาย N
วิธีการของ F. Sanger l U 1945 r. นักชีวเคมีชาวอังกฤษ F. Sanger ได้พัฒนาวิธีแรกในการระบุกรดอะมิโนที่ปลาย N
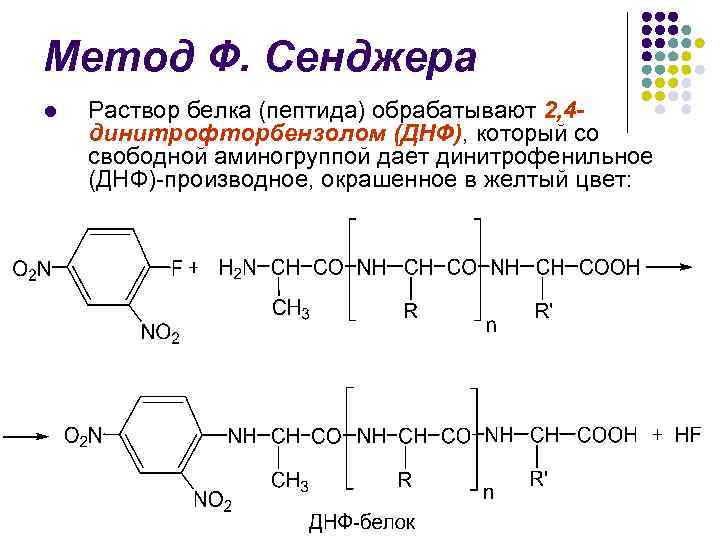 F. วิธีการของแซงเจอร์ l โปรตีน (เปปไทด์) ได้รับการบำบัดด้วย 2, 4 ไดไนโตรฟลูออโรเบนซีน (DNF) ซึ่งจากกลุ่มอะมิโนอิสระจะให้ไดไนโตรฟีนิล (DNF) ซึ่งมีสีเหลือง:
F. วิธีการของแซงเจอร์ l โปรตีน (เปปไทด์) ได้รับการบำบัดด้วย 2, 4 ไดไนโตรฟลูออโรเบนซีน (DNF) ซึ่งจากกลุ่มอะมิโนอิสระจะให้ไดไนโตรฟีนิล (DNF) ซึ่งมีสีเหลือง:
 วิธีการของ F. Sanger l l การไฮโดรไลซิสของกรดที่น่ารังเกียจ (5.7 N HCl) นำไปสู่การทำลายพันธะเปปไทด์และการสร้างกรดอะมิโนที่ปลาย N ที่คล้าย DNP
วิธีการของ F. Sanger l l การไฮโดรไลซิสของกรดที่น่ารังเกียจ (5.7 N HCl) นำไปสู่การทำลายพันธะเปปไทด์และการสร้างกรดอะมิโนที่ปลาย N ที่คล้าย DNP
 วิธี Dansilny l Rozrobleny ในปี 1963 นักชีวเคมีชาวอังกฤษ W. Grey และ B. Hartley
วิธี Dansilny l Rozrobleny ในปี 1963 นักชีวเคมีชาวอังกฤษ W. Grey และ B. Hartley
 วิธี Dansil l ขั้นตอนแรก - ปฏิกิริยาของ dansyl คลอไรด์ (1-dimethylaminonaphthalene-5-sulfochloride) กับเปปไทด์กลุ่มอะมิโนที่ไม่มีการโปรตอน (หรือโปรตีน) กับเปปไทด์ dansyl ดัดแปลง (DNS เปปไทด์)
วิธี Dansil l ขั้นตอนแรก - ปฏิกิริยาของ dansyl คลอไรด์ (1-dimethylaminonaphthalene-5-sulfochloride) กับเปปไทด์กลุ่มอะมิโนที่ไม่มีการโปรตอน (หรือโปรตีน) กับเปปไทด์ dansyl ดัดแปลง (DNS เปปไทด์)

 วิธีที่แข็งแกร่งที่สุดในการกำจัดโปรตีน DNS นั้นอยู่ภายใต้การไฮโดรไลซิสเมื่อมี 5, 7 n HCl ที่ 105 ในช่วงเวลา 12-16 ปี หลังจากนั้นกรด DNS-amino ถูกแยกออก ระบุโดยการเรืองแสงในการแลกเปลี่ยน UV
วิธีที่แข็งแกร่งที่สุดในการกำจัดโปรตีน DNS นั้นอยู่ภายใต้การไฮโดรไลซิสเมื่อมี 5, 7 n HCl ที่ 105 ในช่วงเวลา 12-16 ปี หลังจากนั้นกรด DNS-amino ถูกแยกออก ระบุโดยการเรืองแสงในการแลกเปลี่ยน UV


 วิธี S. Akabori l l เมื่อเปปไทด์ (หรือโปรตีน) ถูกให้ความร้อนด้วยแอนไฮดรัสไฮดราซีนที่อุณหภูมิ 100-120 C พันธะเปปไทด์จะถูกไฮโดรไลซ์ด้วยกรดอะมิโนของไฮดราไซด์ กรดอะมิโนที่ปลาย C จะสูญเสียไปในลักษณะที่ปรากฏ กรดอะมิโนอื่น ๆ สามารถ เห็นได้จากส่วนผสมและระบุได้ว่า:
วิธี S. Akabori l l เมื่อเปปไทด์ (หรือโปรตีน) ถูกให้ความร้อนด้วยแอนไฮดรัสไฮดราซีนที่อุณหภูมิ 100-120 C พันธะเปปไทด์จะถูกไฮโดรไลซ์ด้วยกรดอะมิโนของไฮดราไซด์ กรดอะมิโนที่ปลาย C จะสูญเสียไปในลักษณะที่ปรากฏ กรดอะมิโนอื่น ๆ สามารถ เห็นได้จากส่วนผสมและระบุได้ว่า:

 วิธีการของ P. Edman l วิธีการย่อยสลายโพลีเปปไทด์มีดหมอด้วยความช่วยเหลือของฟีนิลไอโซไทโอเนต (FITC) ถูกย่อยสลายโดยนักเคมีชาวสวีเดน P. V. Edman ในปี 1950-56 l วิธี Edman ช่วยให้สามารถกำจัดกรดอะมิโนที่ตกค้างใน N-terminal ออกจากรูปแบบของ phenylthiohydantoins (PTH) ได้อย่างสม่ำเสมอ l วงจรการเสื่อมสภาพของผิวหนังประกอบด้วย 3 ขั้นตอน
วิธีการของ P. Edman l วิธีการย่อยสลายโพลีเปปไทด์มีดหมอด้วยความช่วยเหลือของฟีนิลไอโซไทโอเนต (FITC) ถูกย่อยสลายโดยนักเคมีชาวสวีเดน P. V. Edman ในปี 1950-56 l วิธี Edman ช่วยให้สามารถกำจัดกรดอะมิโนที่ตกค้างใน N-terminal ออกจากรูปแบบของ phenylthiohydantoins (PTH) ได้อย่างสม่ำเสมอ l วงจรการเสื่อมสภาพของผิวหนังประกอบด้วย 3 ขั้นตอน
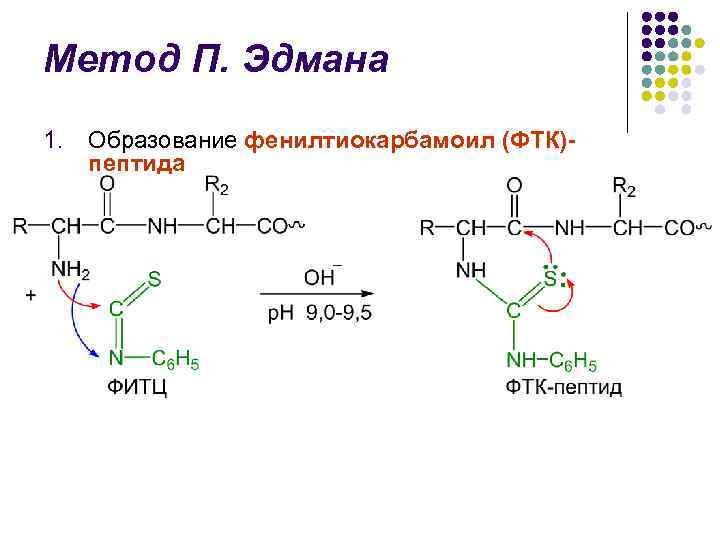



 วิธีการของ P. Edman l l l l การระบุการแยก PTG นั้นเริ่มแรกในวิธี Edman ในช่วงสามชั่วโมงที่ผ่านมามีการดำเนินการโครมาโตกราฟีบนกระดาษจากนั้นจึงใช้โครมาโตกราฟีทรงกลมขนาดเล็กบนซิลิกาเจลและโพลีเอไมด์ซึ่งเป็นโครมาโตกราฟีดั้งเดิมของก๊าซธรรมชาติ วิจัย. เครื่องหาลำดับ Edman และ J. Begg (จากลำดับภาษาอังกฤษ - ลำดับ) - อุปกรณ์ที่มีประสิทธิภาพสูง แยกกรดอะมิโนที่ตกค้างที่ปลาย N โดยอัตโนมัติโดยใช้วิธี Edman
วิธีการของ P. Edman l l l l การระบุการแยก PTG นั้นเริ่มแรกในวิธี Edman ในช่วงสามชั่วโมงที่ผ่านมามีการดำเนินการโครมาโตกราฟีบนกระดาษจากนั้นจึงใช้โครมาโตกราฟีทรงกลมขนาดเล็กบนซิลิกาเจลและโพลีเอไมด์ซึ่งเป็นโครมาโตกราฟีดั้งเดิมของก๊าซธรรมชาติ วิจัย. เครื่องหาลำดับ Edman และ J. Begg (จากลำดับภาษาอังกฤษ - ลำดับ) - อุปกรณ์ที่มีประสิทธิภาพสูง แยกกรดอะมิโนที่ตกค้างที่ปลาย N โดยอัตโนมัติโดยใช้วิธี Edman
 l เพื่อตรวจสอบโครงสร้างของเปปไทด์และโปรตีน สามารถย่อสิ่งต่อไปนี้ได้: l l เอนไซม์ที่กระตุ้นการกำจัดกรดอะมิโนที่ตกค้างที่ปลาย N และ C - อะมิโนคาร์บอกซีเพปทิเดส, วิธีเคมีกายภาพ, สเปกโทรสโกปี, แมสสเปกโตรเมทรี yu
l เพื่อตรวจสอบโครงสร้างของเปปไทด์และโปรตีน สามารถย่อสิ่งต่อไปนี้ได้: l l เอนไซม์ที่กระตุ้นการกำจัดกรดอะมิโนที่ตกค้างที่ปลาย N และ C - อะมิโนคาร์บอกซีเพปทิเดส, วิธีเคมีกายภาพ, สเปกโทรสโกปี, แมสสเปกโตรเมทรี yu
 การวิเคราะห์ข้อมูลเกี่ยวกับโครงสร้างหลักทำให้สามารถพัฒนาหลักการพื้นฐานเพิ่มเติมได้ ความเสถียรของโครงสร้างหลักนั้นมั่นใจได้ด้วยพันธะเปปไทด์เป็นหลัก การมีส่วนร่วมที่เป็นไปได้และพันธะไดซัลไฟด์จำนวนเล็กน้อย ใน polypeptide lance สามารถตรวจพบการรวมกันของกรดอะมิโนที่แตกต่างกัน ใน polypeptides dipeptides ที่เป็นไปได้ทั้งหมดจะถูกเปิดเผย
การวิเคราะห์ข้อมูลเกี่ยวกับโครงสร้างหลักทำให้สามารถพัฒนาหลักการพื้นฐานเพิ่มเติมได้ ความเสถียรของโครงสร้างหลักนั้นมั่นใจได้ด้วยพันธะเปปไทด์เป็นหลัก การมีส่วนร่วมที่เป็นไปได้และพันธะไดซัลไฟด์จำนวนเล็กน้อย ใน polypeptide lance สามารถตรวจพบการรวมกันของกรดอะมิโนที่แตกต่างกัน ใน polypeptides dipeptides ที่เป็นไปได้ทั้งหมดจะถูกเปิดเผย
 การวิเคราะห์ข้อมูลเกี่ยวกับโครงสร้างหลักช่วยให้เราสามารถพัฒนาหลักการพื้นฐานดังต่อไปนี้: l l ผิวหนังของโปรตีนที่เป็นเนื้อเดียวกันแต่ละชนิดมีลักษณะเฉพาะด้วยโครงสร้างหลักที่มีเอกลักษณ์; บ่อยครั้งที่การเปลี่ยนกรดอะมิโนไม่เพียงแต่นำไปสู่การเปลี่ยนแปลงโครงสร้างเท่านั้น แต่ยังรวมถึงการเปลี่ยนแปลงคุณสมบัติทางกายภาพและเคมี และการทำงานทางชีววิทยาด้วย หลักการของความคล้ายคลึงกันของโครงสร้างนี้เป็นเรื่องปกติมากที่สุดสำหรับเอนไซม์โปรตีโอไลติกต่ำ - ทริปซิน, ไคโมทริปซิน ฯลฯ
การวิเคราะห์ข้อมูลเกี่ยวกับโครงสร้างหลักช่วยให้เราสามารถพัฒนาหลักการพื้นฐานดังต่อไปนี้: l l ผิวหนังของโปรตีนที่เป็นเนื้อเดียวกันแต่ละชนิดมีลักษณะเฉพาะด้วยโครงสร้างหลักที่มีเอกลักษณ์; บ่อยครั้งที่การเปลี่ยนกรดอะมิโนไม่เพียงแต่นำไปสู่การเปลี่ยนแปลงโครงสร้างเท่านั้น แต่ยังรวมถึงการเปลี่ยนแปลงคุณสมบัติทางกายภาพและเคมี และการทำงานทางชีววิทยาด้วย หลักการของความคล้ายคลึงกันของโครงสร้างนี้เป็นเรื่องปกติมากที่สุดสำหรับเอนไซม์โปรตีโอไลติกต่ำ - ทริปซิน, ไคโมทริปซิน ฯลฯ

 ในการหลอมรวมแบบไม่มีขั้ว พลังงานของเอ็นน้ำ –CO NNN– จะเข้าใกล้ 16.7 kJ/mol และการเปลี่ยนแปลงขั้วของกระแสน้ำกลางน้ำจะลดพลังงานนี้ลง
ในการหลอมรวมแบบไม่มีขั้ว พลังงานของเอ็นน้ำ –CO NNN– จะเข้าใกล้ 16.7 kJ/mol และการเปลี่ยนแปลงขั้วของกระแสน้ำกลางน้ำจะลดพลังงานนี้ลง
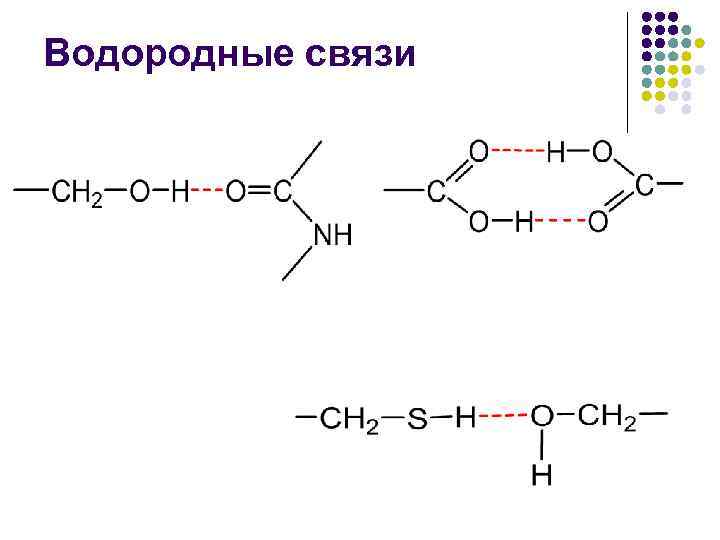
 ปฏิกิริยาที่ไม่ชอบน้ำมีลักษณะเอนโทรปิกเนื่องจากการที่ตัวป้องกันที่ไม่มีขั้วลงไปในน้ำและพยายามสร้างการสัมผัสกับน้ำ ตัวอย่างเช่น น้ำไม่ได้เปลี่ยนโครงสร้างโครงสร้างใหม่และราวกับว่าเป็นกลุ่มพรีมัส คลัสเตอร์ที่มีพลังงานน้อยที่สุด ส่วนเกิน
ปฏิกิริยาที่ไม่ชอบน้ำมีลักษณะเอนโทรปิกเนื่องจากการที่ตัวป้องกันที่ไม่มีขั้วลงไปในน้ำและพยายามสร้างการสัมผัสกับน้ำ ตัวอย่างเช่น น้ำไม่ได้เปลี่ยนโครงสร้างโครงสร้างใหม่และราวกับว่าเป็นกลุ่มพรีมัส คลัสเตอร์ที่มีพลังงานน้อยที่สุด ส่วนเกิน

 ปฏิสัมพันธ์ของ Van der Waals l l ประกอบด้วยแรงกระจาย แรงโน้มถ่วงของอะตอม และแรงในการปรับตัวร่วมกันของเปลือกอิเล็กทรอนิกส์ พลังงานที่ส่งผลต่อการสัมผัสทางผิวหนังมีน้อย (
ปฏิสัมพันธ์ของ Van der Waals l l ประกอบด้วยแรงกระจาย แรงโน้มถ่วงของอะตอม และแรงในการปรับตัวร่วมกันของเปลือกอิเล็กทรอนิกส์ พลังงานที่ส่งผลต่อการสัมผัสทางผิวหนังมีน้อย (
 อันตรกิริยาของไอออนิกหรือไฟฟ้าสถิต l อันตรกิริยาของกลุ่มประจุ l l อันตรกิริยาของหมู่ไอออนิกที่พันธะเกลือสร้างขึ้น พลังงานของพันธะเกลือสามารถเข้าถึงพื้นผิวที่ไม่ชอบน้ำ 41.9 k. J/mol แต่ปริมาณของพวกมันค่อนข้างน้อย ปฏิกิริยาระหว่างไอออน - ไดโพลและไดโพล - ไดโพลที่คล้ายกัน
อันตรกิริยาของไอออนิกหรือไฟฟ้าสถิต l อันตรกิริยาของกลุ่มประจุ l l อันตรกิริยาของหมู่ไอออนิกที่พันธะเกลือสร้างขึ้น พลังงานของพันธะเกลือสามารถเข้าถึงพื้นผิวที่ไม่ชอบน้ำ 41.9 k. J/mol แต่ปริมาณของพวกมันค่อนข้างน้อย ปฏิกิริยาระหว่างไอออน - ไดโพลและไดโพล - ไดโพลที่คล้ายกัน
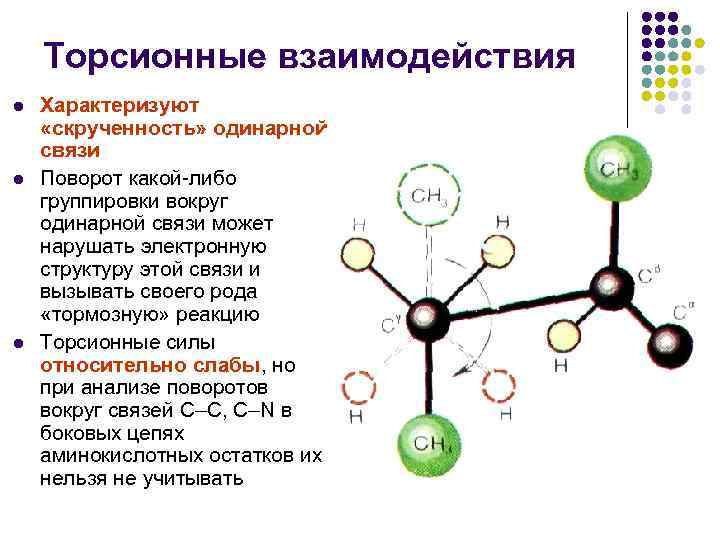 ปฏิกิริยาระหว่างแรงบิด l l l l แสดงลักษณะของ "การบิด" ของพันธะเดี่ยว การหมุนของการรวมกลุ่มใดๆ รอบพันธะเดี่ยวสามารถทำลายโครงสร้างทางอิเล็กทรอนิกส์ของพันธะและกระตุ้นให้เกิดปฏิกิริยา "halm" ปฏิกิริยาแรงบิดและแรงจะอ่อนแออย่างเห็นได้ชัด แต่เมื่อ การวิเคราะห์การหมุนรอบเอ็น С–С, С–N ของเสีย มีดหมอ กรดอะมิโนส่วนเกินไม่สามารถหลีกเลี่ยงได้
ปฏิกิริยาระหว่างแรงบิด l l l l แสดงลักษณะของ "การบิด" ของพันธะเดี่ยว การหมุนของการรวมกลุ่มใดๆ รอบพันธะเดี่ยวสามารถทำลายโครงสร้างทางอิเล็กทรอนิกส์ของพันธะและกระตุ้นให้เกิดปฏิกิริยา "halm" ปฏิกิริยาแรงบิดและแรงจะอ่อนแออย่างเห็นได้ชัด แต่เมื่อ การวิเคราะห์การหมุนรอบเอ็น С–С, С–N ของเสีย มีดหมอ กรดอะมิโนส่วนเกินไม่สามารถหลีกเลี่ยงได้

 โครงสร้างรองของโปรตีน... l l l การขยายตัวอย่างกว้างขวางของส่วนโค้งมนของ polypeptide lancet โดยไม่มีชนิดและโครงสร้างของอนุมูลของกรดอะมิโน มันถูกสร้างขึ้นโดยปฏิสัมพันธ์ของพันธะน้ำระหว่างกลุ่มเปปไทด์ของทั้ง lancet เดียวและต่างกัน . ลันต์สยูกิฟ
โครงสร้างรองของโปรตีน... l l l การขยายตัวอย่างกว้างขวางของส่วนโค้งมนของ polypeptide lancet โดยไม่มีชนิดและโครงสร้างของอนุมูลของกรดอะมิโน มันถูกสร้างขึ้นโดยปฏิสัมพันธ์ของพันธะน้ำระหว่างกลุ่มเปปไทด์ของทั้ง lancet เดียวและต่างกัน . ลันต์สยูกิฟ
 โครงสร้างทุติยภูมิของโปรตีน l l l ทุกส่วนของโมเลกุลโปรตีนมีโครงสร้างรอง บางคนพิจารณาโครงสร้างรองขององค์ประกอบที่มีคาบมากกว่า: - เกลียวและ - โครงสร้าง อย่างไรก็ตาม ในโปรตีน ส่วนต่างๆ ซึ่งมีโครงสร้างจะมีความแม่นยำมากขึ้นใน วิธีที่อ่อนโยนแม้ว่าโครงสร้างที่กว้างขวางของมันจะไม่ได้ล้างแค้นใครก็ตามเป็นครั้งคราวซึ่งเป็นบรรทัดฐานปกติ ก่อนหน้านั้น เราเข้าใจแนวคิดของโครงสร้างทุติยภูมิอย่างถ่องแท้ โครงสร้างทุติยภูมิมี 2 ประเภท: แบบปกติและไม่สม่ำเสมอ แนวคิดของโครงสร้างทุติยภูมิใช้ไม่ได้กับโมเลกุลโปรตีนทั้งหมดโดยรวม แต่ใช้กับส่วนที่ใหญ่กว่าเล็กน้อยของพื้น มีดหมอไอเปปไทด์
โครงสร้างทุติยภูมิของโปรตีน l l l ทุกส่วนของโมเลกุลโปรตีนมีโครงสร้างรอง บางคนพิจารณาโครงสร้างรองขององค์ประกอบที่มีคาบมากกว่า: - เกลียวและ - โครงสร้าง อย่างไรก็ตาม ในโปรตีน ส่วนต่างๆ ซึ่งมีโครงสร้างจะมีความแม่นยำมากขึ้นใน วิธีที่อ่อนโยนแม้ว่าโครงสร้างที่กว้างขวางของมันจะไม่ได้ล้างแค้นใครก็ตามเป็นครั้งคราวซึ่งเป็นบรรทัดฐานปกติ ก่อนหน้านั้น เราเข้าใจแนวคิดของโครงสร้างทุติยภูมิอย่างถ่องแท้ โครงสร้างทุติยภูมิมี 2 ประเภท: แบบปกติและไม่สม่ำเสมอ แนวคิดของโครงสร้างทุติยภูมิใช้ไม่ได้กับโมเลกุลโปรตีนทั้งหมดโดยรวม แต่ใช้กับส่วนที่ใหญ่กว่าเล็กน้อยของพื้น มีดหมอไอเปปไทด์
 โครงสร้างทุติยภูมิของโปรตีน ปฏิกิริยาที่มีบทบาทที่สำคัญที่สุดในโครงสร้างทุติยภูมิที่เกิดขึ้นจะถูกระบุโดย: ล. ลักษณะของพันธะเปปไทด์และปฏิกิริยาแบบสเตอริก (- และ - ตัด) บทบาทหลักแสดงโดยพันธะน้ำระหว่าง กลุ่มเปปไทด์ ซึ่งทำซ้ำเป็นระยะใน Lancus
โครงสร้างทุติยภูมิของโปรตีน ปฏิกิริยาที่มีบทบาทที่สำคัญที่สุดในโครงสร้างทุติยภูมิที่เกิดขึ้นจะถูกระบุโดย: ล. ลักษณะของพันธะเปปไทด์และปฏิกิริยาแบบสเตอริก (- และ - ตัด) บทบาทหลักแสดงโดยพันธะน้ำระหว่าง กลุ่มเปปไทด์ ซึ่งทำซ้ำเป็นระยะใน Lancus
 การรักษาเสถียรภาพของโครงสร้างรองสำหรับการก่อตัวของสารยึดเกาะน้ำ l l การฉีดโปรตีนส่วนเกินด้วยน้ำ น้ำสามารถแข่งขันกับการก่อตัวของสารยึดเกาะน้ำได้อย่างมีประสิทธิภาพ: การแช่น้ำจะลดลงเมื่อสร้างโครงสร้างโปรตีนที่มีขนาดกะทัดรัด การเจริญเติบโต และแทนที่พันธะเปปไทด์ เพิ่มความเข้ากันได้ของการโต้ตอบ ดังนั้น โครงสร้างรองที่มีความเสถียรจึงอยู่ในโครงสร้างตติยภูมิที่มีขนาดกะทัดรัด
การรักษาเสถียรภาพของโครงสร้างรองสำหรับการก่อตัวของสารยึดเกาะน้ำ l l การฉีดโปรตีนส่วนเกินด้วยน้ำ น้ำสามารถแข่งขันกับการก่อตัวของสารยึดเกาะน้ำได้อย่างมีประสิทธิภาพ: การแช่น้ำจะลดลงเมื่อสร้างโครงสร้างโปรตีนที่มีขนาดกะทัดรัด การเจริญเติบโต และแทนที่พันธะเปปไทด์ เพิ่มความเข้ากันได้ของการโต้ตอบ ดังนั้น โครงสร้างรองที่มีความเสถียรจึงอยู่ในโครงสร้างตติยภูมิที่มีขนาดกะทัดรัด

 -Spiral l ในหินยุค 50 ของศตวรรษที่ XX L. Pauling และ R. Corey จากข้อมูลเกี่ยวกับโครงสร้างของผลึกของกรดอะมิโนและเปปไทด์อย่างง่ายได้พิจารณาถึงความสอดคล้องเป็นระยะ ๆ ที่เป็นไปได้ของ polypeptide lanjug และได้ข้อสรุปว่าโครงสร้างที่เป็นเนื้อเดียวกันมากที่สุดนั้นถูกตั้งชื่อโดยพวกเขา -spi rally
-Spiral l ในหินยุค 50 ของศตวรรษที่ XX L. Pauling และ R. Corey จากข้อมูลเกี่ยวกับโครงสร้างของผลึกของกรดอะมิโนและเปปไทด์อย่างง่ายได้พิจารณาถึงความสอดคล้องเป็นระยะ ๆ ที่เป็นไปได้ของ polypeptide lanjug และได้ข้อสรุปว่าโครงสร้างที่เป็นเนื้อเดียวกันมากที่สุดนั้นถูกตั้งชื่อโดยพวกเขา -spi rally
 -เกลียว การเลือกตำแหน่งจะขึ้นอยู่กับเกณฑ์ต่อไปนี้: 1. การสร้างโครงสร้างที่อัดแน่นแน่นโดยไม่ว่างเปล่าและวิพากษ์วิจารณ์อะตอมมากเกินไป 2. ความอิ่มตัวสูงสุดของโครงสร้างด้วยพันธะน้ำจากสารเคมีนี้ ดังนั้นเรขาคณิตจึงใกล้เคียงกับเส้นตรง 3 . Dotrimannya ระหว่างอะตอมไรเซอร์และมุมและให้อภัยเปปไทด์ l
-เกลียว การเลือกตำแหน่งจะขึ้นอยู่กับเกณฑ์ต่อไปนี้: 1. การสร้างโครงสร้างที่อัดแน่นแน่นโดยไม่ว่างเปล่าและวิพากษ์วิจารณ์อะตอมมากเกินไป 2. ความอิ่มตัวสูงสุดของโครงสร้างด้วยพันธะน้ำจากสารเคมีนี้ ดังนั้นเรขาคณิตจึงใกล้เคียงกับเส้นตรง 3 . Dotrimannya ระหว่างอะตอมไรเซอร์และมุมและให้อภัยเปปไทด์ l
 -เกลียว l ในจิตใจของคนเหล่านี้อาจมีทั้งเกลียวขวาและเกลียวซ้าย แต่เกลียวขวาจะดูโดดเด่นกว่าด้านซ้ายเนื่องจากเปปไทด์แลนซ์ถูกสร้างขึ้นด้วยกรดแอล-อะมิโน
-เกลียว l ในจิตใจของคนเหล่านี้อาจมีทั้งเกลียวขวาและเกลียวซ้าย แต่เกลียวขวาจะดูโดดเด่นกว่าด้านซ้ายเนื่องจากเปปไทด์แลนซ์ถูกสร้างขึ้นด้วยกรดแอล-อะมิโน
 -เกลียว l l โครงสร้างขดลวดพัฒนาขึ้นเมื่อทุกด้านของมีดหมอโพลีเปปไทด์หมุนไปรอบเอ็นเชิงเดี่ยว (i) มีค่าและเครื่องหมายเท่ากัน (ใกล้ถึง 60) ซึ่งนำไปสู่การบิดมีดหมอทีละขั้นตอน ณ จุดใด อนุมูลมีค่าไม่มีกรดมากเกินไปปรากฏที่ขอบของเกลียวทรงกระบอกที่ละลายและสามารถตรวจสอบให้แน่ใจว่าลักษณะที่ไม่ชอบน้ำหรือชอบน้ำของพื้นผิวทรงกระบอกขึ้นอยู่กับลักษณะของพวกมัน
-เกลียว l l โครงสร้างขดลวดพัฒนาขึ้นเมื่อทุกด้านของมีดหมอโพลีเปปไทด์หมุนไปรอบเอ็นเชิงเดี่ยว (i) มีค่าและเครื่องหมายเท่ากัน (ใกล้ถึง 60) ซึ่งนำไปสู่การบิดมีดหมอทีละขั้นตอน ณ จุดใด อนุมูลมีค่าไม่มีกรดมากเกินไปปรากฏที่ขอบของเกลียวทรงกระบอกที่ละลายและสามารถตรวจสอบให้แน่ใจว่าลักษณะที่ไม่ชอบน้ำหรือชอบน้ำของพื้นผิวทรงกระบอกขึ้นอยู่กับลักษณะของพวกมัน
 - พารามิเตอร์ทางเรขาคณิตของเกลียว: l l l l รัศมี r = 2.3 Å (0.23 nm) ความสูงของเกลียว (การกระจัด) ต่อส่วนเกิน 1 d = 1.5 Å ขอบของเกลียว (ช่วงการระบุตัวตน) P = 5.4 Å 1 รอบของเกลียว รูปร่าง กรดอะมิโน 3, 6 ส่วนเกินทั้งหมด – พันธะ C=O ตรงไปข้างหน้าถึงปลาย C และหมู่ –N–H จะอยู่ด้านหลังตรง ในเกลียว กลุ่มผิวหนัง –NH เชื่อมต่อกันด้วยสายน้ำ ไปยังกลุ่ม –CO ของกรดอะมิโนตัวที่ 4 มากเกินไป znyogo (5 1 โทร)
- พารามิเตอร์ทางเรขาคณิตของเกลียว: l l l l รัศมี r = 2.3 Å (0.23 nm) ความสูงของเกลียว (การกระจัด) ต่อส่วนเกิน 1 d = 1.5 Å ขอบของเกลียว (ช่วงการระบุตัวตน) P = 5.4 Å 1 รอบของเกลียว รูปร่าง กรดอะมิโน 3, 6 ส่วนเกินทั้งหมด – พันธะ C=O ตรงไปข้างหน้าถึงปลาย C และหมู่ –N–H จะอยู่ด้านหลังตรง ในเกลียว กลุ่มผิวหนัง –NH เชื่อมต่อกันด้วยสายน้ำ ไปยังกลุ่ม –CO ของกรดอะมิโนตัวที่ 4 มากเกินไป znyogo (5 1 โทร)
 -Spiral l l จำนวนส่วนของ -spiral ในโปรตีนทรงกลมมีขนาดเล็กมาก (มีกรดอะมิโน 5 - 15 ส่วนเกิน, 3 - 4 รอบของเกลียว) ในโปรตีนไฟบริลลารี - ขยายออกอย่างมาก กรุณาโทรติดต่อในสถานที่ที่มีการเปิดส่วนเกิน เพื่อให้ระบบสามารถถูกรบกวนได้ ณ จุดนี้เกลียวทั้งหมดหดตัวที่ 20-30
-Spiral l l จำนวนส่วนของ -spiral ในโปรตีนทรงกลมมีขนาดเล็กมาก (มีกรดอะมิโน 5 - 15 ส่วนเกิน, 3 - 4 รอบของเกลียว) ในโปรตีนไฟบริลลารี - ขยายออกอย่างมาก กรุณาโทรติดต่อในสถานที่ที่มีการเปิดส่วนเกิน เพื่อให้ระบบสามารถถูกรบกวนได้ ณ จุดนี้เกลียวทั้งหมดหดตัวที่ 20-30

 - โครงสร้าง l l l เสนอโดย W. T. Astbury ในปี 1941 บนพื้นฐานของการศึกษาโครงสร้างเอ็กซ์เรย์ - เคราติน หลังจากผ่านไป 10 ปี L. Pauling และ R. Corey ได้กำหนดว่าโครงสร้างหรือ "แผ่นพับ" นั้นมีสาเหตุมาจากการรักษาเสถียรภาพของเอ็นน้ำระหว่าง lancinar ที่เชื่อมโยงการถักนิตติ้งซิกแซก เอ็นเปปไทด์
- โครงสร้าง l l l เสนอโดย W. T. Astbury ในปี 1941 บนพื้นฐานของการศึกษาโครงสร้างเอ็กซ์เรย์ - เคราติน หลังจากผ่านไป 10 ปี L. Pauling และ R. Corey ได้กำหนดว่าโครงสร้างหรือ "แผ่นพับ" นั้นมีสาเหตุมาจากการรักษาเสถียรภาพของเอ็นน้ำระหว่าง lancinar ที่เชื่อมโยงการถักนิตติ้งซิกแซก เอ็นเปปไทด์




 -โครงสร้าง l l จำนวนกรดอะมิโนส่วนเกินในส่วนของเพปไทด์ lancelet ซึ่งสร้างโครงสร้าง มีตั้งแต่ 3 ถึง 8 โครงสร้างที่ขยายออก เรียกว่าลูกบอลหรือแผ่นพับ มักประกอบด้วย 2- 6 lancelets บางครั้งมากถึง 10
-โครงสร้าง l l จำนวนกรดอะมิโนส่วนเกินในส่วนของเพปไทด์ lancelet ซึ่งสร้างโครงสร้าง มีตั้งแต่ 3 ถึง 8 โครงสร้างที่ขยายออก เรียกว่าลูกบอลหรือแผ่นพับ มักประกอบด้วย 2- 6 lancelets บางครั้งมากถึง 10
 โครงสร้าง l l l l เมื่อโครงสร้างถูกสร้างขึ้น กลุ่มที่เป็นกรดของกรดอะมิโนส่วนเกินจะถูกเปิดเผยโดยพื้นผิวด้านต่างๆ พื้นผิวนั้นมีรูปร่างพับ และรอยพับถูกกำหนดโดยอะตอมของคาร์บอน กลุ่มอินทรีย์ซึ่ง มาจากพวกมัน ถูกสร้างขึ้น หวี ซึ่งช่วยให้คุณสามารถปั้นและขยายพื้นผิวยาวที่เต็มไปด้วยอนุมูลทางชีวภาพประเภทเดียวกัน (เช่น ไม่ชอบน้ำ) ของพื้นผิวที่ไม่ชอบน้ำของลูกบอลพับโต้ตอบกันหรือกับสันเขาที่ไม่ชอบน้ำ เป็นส่วนหนึ่งของนิวเคลียสที่ไม่ชอบน้ำโมเลกุลภายในซึ่งทำให้โครงสร้างของโปรตีนมีความเสถียรได้ง่าย
โครงสร้าง l l l l เมื่อโครงสร้างถูกสร้างขึ้น กลุ่มที่เป็นกรดของกรดอะมิโนส่วนเกินจะถูกเปิดเผยโดยพื้นผิวด้านต่างๆ พื้นผิวนั้นมีรูปร่างพับ และรอยพับถูกกำหนดโดยอะตอมของคาร์บอน กลุ่มอินทรีย์ซึ่ง มาจากพวกมัน ถูกสร้างขึ้น หวี ซึ่งช่วยให้คุณสามารถปั้นและขยายพื้นผิวยาวที่เต็มไปด้วยอนุมูลทางชีวภาพประเภทเดียวกัน (เช่น ไม่ชอบน้ำ) ของพื้นผิวที่ไม่ชอบน้ำของลูกบอลพับโต้ตอบกันหรือกับสันเขาที่ไม่ชอบน้ำ เป็นส่วนหนึ่งของนิวเคลียสที่ไม่ชอบน้ำโมเลกุลภายในซึ่งทำให้โครงสร้างของโปรตีนมีความเสถียรได้ง่าย

 -Vigin l l ทั้งเกลียวและโครงสร้างแสดงอยู่ในโปรตีนทรงกลมในส่วนสั้น ๆ ซึ่งหมายความว่าส่วนหนึ่งของโครงสร้างรองของโปรตีนตกอยู่บนลูปประเภทต่าง ๆ ซึ่งช่วยให้คุณเปลี่ยนทิศทางของเปปไทด์ lanjug ได้ แบบประหยัด องค์ประกอบโครงสร้างที่ทำให้โพลีเปปไทด์สามารถหมุนได้ 180 องศา การแบ่งกลุ่มเปปไทด์ 3 กลุ่ม - -vigin เสถียรด้วยพันธะน้ำเดียว มักปรากฏบนพื้นผิวของโปรตีนโกลบูลซึ่งมักจะมีบทบาทสำคัญในการทำปฏิกิริยากับโมเลกุลอื่น ๆ
-Vigin l l ทั้งเกลียวและโครงสร้างแสดงอยู่ในโปรตีนทรงกลมในส่วนสั้น ๆ ซึ่งหมายความว่าส่วนหนึ่งของโครงสร้างรองของโปรตีนตกอยู่บนลูปประเภทต่าง ๆ ซึ่งช่วยให้คุณเปลี่ยนทิศทางของเปปไทด์ lanjug ได้ แบบประหยัด องค์ประกอบโครงสร้างที่ทำให้โพลีเปปไทด์สามารถหมุนได้ 180 องศา การแบ่งกลุ่มเปปไทด์ 3 กลุ่ม - -vigin เสถียรด้วยพันธะน้ำเดียว มักปรากฏบนพื้นผิวของโปรตีนโกลบูลซึ่งมักจะมีบทบาทสำคัญในการทำปฏิกิริยากับโมเลกุลอื่น ๆ


 l ผลการวิเคราะห์โครงสร้างด้วยรังสีเอกซ์พบว่าวิธีการพับโมเลกุลโปรตีนนั้นถูกกำหนดโดยลำดับกรดอะมิโนของมัน - gly, pro, asn Yakshcho โดยมีกรดอะมิโนส่วนเกิน 6 กลุ่ม 4 รวมการสร้างเกลียวแล้วกลุ่มนี้ เป็นศูนย์กลางของเกลียว หากนำ 3 สิ่งพิเศษจาก 5 กลุ่มมารวมกันเป็นโครงสร้าง การจัดกลุ่มนี้ จะเป็นเมล็ดพันธุ์สำหรับ
l ผลการวิเคราะห์โครงสร้างด้วยรังสีเอกซ์พบว่าวิธีการพับโมเลกุลโปรตีนนั้นถูกกำหนดโดยลำดับกรดอะมิโนของมัน - gly, pro, asn Yakshcho โดยมีกรดอะมิโนส่วนเกิน 6 กลุ่ม 4 รวมการสร้างเกลียวแล้วกลุ่มนี้ เป็นศูนย์กลางของเกลียว หากนำ 3 สิ่งพิเศษจาก 5 กลุ่มมารวมกันเป็นโครงสร้าง การจัดกลุ่มนี้ จะเป็นเมล็ดพันธุ์สำหรับ

 โครงสร้างรอง (เหนือรอง) ของโปรตีน l l ระดับของการจัดระเบียบของโมเลกุลโปรตีนนี้เกี่ยวข้องกับการมีอยู่ของโครงสร้างรองที่มีปฏิสัมพันธ์ซึ่งกันและกัน
โครงสร้างรอง (เหนือรอง) ของโปรตีน l l ระดับของการจัดระเบียบของโมเลกุลโปรตีนนี้เกี่ยวข้องกับการมีอยู่ของโครงสร้างรองที่มีปฏิสัมพันธ์ซึ่งกันและกัน

 Superspiralized -spiral l l เกิดจากการบิดรูปลอก -เกลียวที่โปรโตไฟบริล (-เคราติน) ซึ่งรวมตัวเป็นไมโครไฟบริล -เกลียวถูกกดเข้าด้วยกันโดยแรงของ Van der Waals
Superspiralized -spiral l l เกิดจากการบิดรูปลอก -เกลียวที่โปรโตไฟบริล (-เคราติน) ซึ่งรวมตัวเป็นไมโครไฟบริล -เกลียวถูกกดเข้าด้วยกันโดยแรงของ Van der Waals

 x l สายรัดซึ่งประกอบด้วยลูกบอลสองลูกขนานกันโดยมีข้อต่อระหว่างพวกมันดูเหมือน: l ลูกบอลที่ไม่เป็นระเบียบ - ด้วย
x l สายรัดซึ่งประกอบด้วยลูกบอลสองลูกขนานกันโดยมีข้อต่อระหว่างพวกมันดูเหมือน: l ลูกบอลที่ไม่เป็นระเบียบ - ด้วย
 x l สายรัดซึ่งประกอบด้วยลูกบอลสองลูกขนานกันโดยมีข้อต่อระหว่างกันดูเหมือนว่า: l l - เกลียว - มีดหมอวางตาม Rossman แผนการที่เชื่อมต่อกันสองแห่งต่อเนื่องกันสร้างโครงสร้างของ "เครื่องประดับกรีก" -
x l สายรัดซึ่งประกอบด้วยลูกบอลสองลูกขนานกันโดยมีข้อต่อระหว่างกันดูเหมือนว่า: l l - เกลียว - มีดหมอวางตาม Rossman แผนการที่เชื่อมต่อกันสองแห่งต่อเนื่องกันสร้างโครงสร้างของ "เครื่องประดับกรีก" -
 x l สายรัดซึ่งประกอบด้วยลูกบอลสองลูกขนานกันโดยมีข้อต่อระหว่างกันดูเหมือนว่า: l -โครงสร้าง - - - ซิกแซก "กุญแจวอลนัท"
x l สายรัดซึ่งประกอบด้วยลูกบอลสองลูกขนานกันโดยมีข้อต่อระหว่างกันดูเหมือนว่า: l -โครงสร้าง - - - ซิกแซก "กุญแจวอลนัท"

 โดเมนและส่วนสั้น ๆ ทีละส่วนของโพลีเปปไทด์แลนซ์ซึ่งเรียกว่าส่วนบานพับโดเมนการทำงานสามารถเกิดขึ้นได้จากโดเมนโครงสร้างหนึ่งหรือหลายโดเมน ในเอนไซม์จำนวนหนึ่ง Active center จะถูกแยกระหว่างโดเมน
โดเมนและส่วนสั้น ๆ ทีละส่วนของโพลีเปปไทด์แลนซ์ซึ่งเรียกว่าส่วนบานพับโดเมนการทำงานสามารถเกิดขึ้นได้จากโดเมนโครงสร้างหนึ่งหรือหลายโดเมน ในเอนไซม์จำนวนหนึ่ง Active center จะถูกแยกระหว่างโดเมน



 3. + -โปรตีนเป็นส่วนต่างๆ ซึ่งส่วนใหญ่ทำจากเกลียว และส่วนต่างๆ ที่สร้างจากอินซูลินที่มีลูกขนานกัน
3. + -โปรตีนเป็นส่วนต่างๆ ซึ่งส่วนใหญ่ทำจากเกลียว และส่วนต่างๆ ที่สร้างจากอินซูลินที่มีลูกขนานกัน

 4. / -โปรตีน l l -เกลียวและ -โครงสร้างถูกสร้างขึ้นตามเส้นทางของ Lancug โครงสร้างส่วนใหญ่ (ขนาน) ได้รับการแปลเป็นภาษาท้องถิ่นในส่วนกลางของโมเลกุลโดยที่โครงสร้างเหล่านี้โค้งงอเมื่อลักษณะของใบพัด (“ โครงสร้างบิด") สร้าง "ฐาน" ที่เข้มงวด "ซึ่งส่วนอื่น ๆ ของโมเลกุลเชื่อมต่อกัน
4. / -โปรตีน l l -เกลียวและ -โครงสร้างถูกสร้างขึ้นตามเส้นทางของ Lancug โครงสร้างส่วนใหญ่ (ขนาน) ได้รับการแปลเป็นภาษาท้องถิ่นในส่วนกลางของโมเลกุลโดยที่โครงสร้างเหล่านี้โค้งงอเมื่อลักษณะของใบพัด (“ โครงสร้างบิด") สร้าง "ฐาน" ที่เข้มงวด "ซึ่งส่วนอื่น ๆ ของโมเลกุลเชื่อมต่อกัน


 โดเมน l l l ขนาดเฉลี่ยของโดเมนคือ 100 – 150 เพิ่มเติม ซึ่งหมายถึงทรงกลมที่มีเส้นผ่านศูนย์กลางประมาณ 2.5 นาโนเมตร จ. การก่อตัวของโครงสร้างตติยภูมิซึ่งเกิดขึ้นในขั้นตอนสุดท้ายของการก่อตัวของทรงกลม
โดเมน l l l ขนาดเฉลี่ยของโดเมนคือ 100 – 150 เพิ่มเติม ซึ่งหมายถึงทรงกลมที่มีเส้นผ่านศูนย์กลางประมาณ 2.5 นาโนเมตร จ. การก่อตัวของโครงสร้างตติยภูมิซึ่งเกิดขึ้นในขั้นตอนสุดท้ายของการก่อตัวของทรงกลม

 โครงสร้างตติยภูมิของโปรตีน... l l l l แสดงคุณลักษณะของการกระจายอย่างกว้างขวางของส่วนที่เรียงลำดับและอสัณฐานในมีดหมอโพลีเปปไทด์โดยรวมซึ่งเนื่องมาจากอันตรกิริยาของอนุมูลทางชีวภาพและอยู่ในประเภทและโครงสร้างของพวกมัน ดังนั้น โครงสร้างตติยภูมิจึงอธิบาย การจัดเรียงเชิงพื้นที่ของโมเลกุลโปรตีนทั้งหมด เนื่องจากสร้างขึ้นจากสารโพลีเปปไทด์ชนิดเดียวที่เกี่ยวข้องโดยตรงกับรูปร่างของโมเลกุลโปรตีน ซึ่งอาจแตกต่างกันไปตั้งแต่แบบเส้นไปจนถึงแบบเกลียว
โครงสร้างตติยภูมิของโปรตีน... l l l l แสดงคุณลักษณะของการกระจายอย่างกว้างขวางของส่วนที่เรียงลำดับและอสัณฐานในมีดหมอโพลีเปปไทด์โดยรวมซึ่งเนื่องมาจากอันตรกิริยาของอนุมูลทางชีวภาพและอยู่ในประเภทและโครงสร้างของพวกมัน ดังนั้น โครงสร้างตติยภูมิจึงอธิบาย การจัดเรียงเชิงพื้นที่ของโมเลกุลโปรตีนทั้งหมด เนื่องจากสร้างขึ้นจากสารโพลีเปปไทด์ชนิดเดียวที่เกี่ยวข้องโดยตรงกับรูปร่างของโมเลกุลโปรตีน ซึ่งอาจแตกต่างกันไปตั้งแต่แบบเส้นไปจนถึงแบบเกลียว
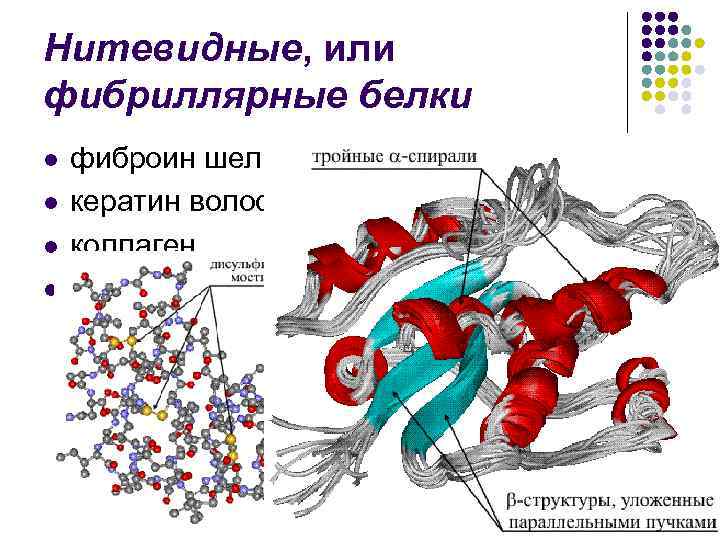 โปรตีนคล้ายไหมหรือไฟบริลลาร์ ไฟโบรอิน เย็บเคราตินของเส้นผม เขา สะสมคอลลาเจน และอื่นๆ
โปรตีนคล้ายไหมหรือไฟบริลลาร์ ไฟโบรอิน เย็บเคราตินของเส้นผม เขา สะสมคอลลาเจน และอื่นๆ

 โครงสร้างระดับตติยภูมิของโปรตีนเป็นพื้นฐานของการทำงานของโปรตีน ซึ่งต้องการการจัดระเบียบเชิงพื้นที่ที่แม่นยำของวงดนตรีขนาดใหญ่ที่สร้างขึ้นโดยไม่มีกรดอะมิโนมากเกินไป จะต้องดำเนินการโปรตีน (การสูญเสียสภาพ) จนกว่าจะสูญเสียความสามารถในการทำงาน
โครงสร้างระดับตติยภูมิของโปรตีนเป็นพื้นฐานของการทำงานของโปรตีน ซึ่งต้องการการจัดระเบียบเชิงพื้นที่ที่แม่นยำของวงดนตรีขนาดใหญ่ที่สร้างขึ้นโดยไม่มีกรดอะมิโนมากเกินไป จะต้องดำเนินการโปรตีน (การสูญเสียสภาพ) จนกว่าจะสูญเสียความสามารถในการทำงาน
 โครงสร้างตติยภูมิของโปรตีน l l l ความเสถียรของโครงสร้างตติยภูมินั้นอยู่ในระบบของปฏิกิริยาที่ไม่ใช่โควาเลนต์ตรงกลางของโปรตีนโกลบูล (?) โปรตีนบางชนิดยังได้รับความเสถียรเพิ่มเติมโดยพันธะโควาเลนต์ - ไดซัลไฟด์ - Glanzug = RTln K = Nchain – T S lanceug Nchain 0 (S
โครงสร้างตติยภูมิของโปรตีน l l l ความเสถียรของโครงสร้างตติยภูมินั้นอยู่ในระบบของปฏิกิริยาที่ไม่ใช่โควาเลนต์ตรงกลางของโปรตีนโกลบูล (?) โปรตีนบางชนิดยังได้รับความเสถียรเพิ่มเติมโดยพันธะโควาเลนต์ - ไดซัลไฟด์ - Glanzug = RTln K = Nchain – T S lanceug Nchain 0 (S
 โครงสร้างระดับตติยภูมิของโปรตีน l Spivvіdnosti Chain i -T Schip อยู่ใน: l l จำนวนของพันธะที่ไม่ใช่โควาเลนต์ที่ติดตั้งเมื่อโมเลกุลโปรตีนถูกกินเข้าไปในทรงกลมและหลังจากโพลีเปปไทด์แลนเซอร์ขึ้นอยู่กับอุณหภูมิ ซี ซรอสทันยัม T |T S| ย้ายเร็วหรือช้า | ยังไม่มี| โครงสร้างแบบเปิดนั้นสูญเสียความเสถียร - การสูญเสียความร้อนของโปรตีน
โครงสร้างระดับตติยภูมิของโปรตีน l Spivvіdnosti Chain i -T Schip อยู่ใน: l l จำนวนของพันธะที่ไม่ใช่โควาเลนต์ที่ติดตั้งเมื่อโมเลกุลโปรตีนถูกกินเข้าไปในทรงกลมและหลังจากโพลีเปปไทด์แลนเซอร์ขึ้นอยู่กับอุณหภูมิ ซี ซรอสทันยัม T |T S| ย้ายเร็วหรือช้า | ยังไม่มี| โครงสร้างแบบเปิดนั้นสูญเสียความเสถียร - การสูญเสียความร้อนของโปรตีน
 โครงสร้างระดับตติยภูมิของโปรตีน l l เมื่อกลืนทรงกลมเอนโทรปีของเปปไทด์แลนซ์จะเปลี่ยนไป แต่ในขณะเดียวกัน เอนโทรปีของผู้ริเริ่มก็เพิ่มขึ้น - น้ำ สิ่งนี้มีบทบาทสำคัญในการรักษาเสถียรภาพของโครงสร้างตติยภูมิ
โครงสร้างระดับตติยภูมิของโปรตีน l l เมื่อกลืนทรงกลมเอนโทรปีของเปปไทด์แลนซ์จะเปลี่ยนไป แต่ในขณะเดียวกัน เอนโทรปีของผู้ริเริ่มก็เพิ่มขึ้น - น้ำ สิ่งนี้มีบทบาทสำคัญในการรักษาเสถียรภาพของโครงสร้างตติยภูมิ
 โครงสร้างระดับตติยภูมิของโปรตีน l l น้ำเป็นสารสร้างโครงสร้างที่ไม่สำคัญและมีลำดับสูง (?) กลุ่มขั้วของโปรตีนสามารถรวมเข้ากับโครงสร้างคล้ายน้ำแข็งของน้ำได้อย่างง่ายดาย โดยพันธะของน้ำจะถูกแทรกเข้าไป กลุ่มที่ไม่มีขั้ว ครอบครองพื้นที่ว่างของตัวเองที่สร้างโดยโมเลกุล น้ำ ผูกด้วยสายน้ำ ทั้งหมดนี้จะปิดการไหลของโมเลกุลของน้ำที่วุ่นวายเพื่อให้มีความเป็นระเบียบการเปลี่ยนแปลงเอนโทรปีของน้ำ
โครงสร้างระดับตติยภูมิของโปรตีน l l น้ำเป็นสารสร้างโครงสร้างที่ไม่สำคัญและมีลำดับสูง (?) กลุ่มขั้วของโปรตีนสามารถรวมเข้ากับโครงสร้างคล้ายน้ำแข็งของน้ำได้อย่างง่ายดาย โดยพันธะของน้ำจะถูกแทรกเข้าไป กลุ่มที่ไม่มีขั้ว ครอบครองพื้นที่ว่างของตัวเองที่สร้างโดยโมเลกุล น้ำ ผูกด้วยสายน้ำ ทั้งหมดนี้จะปิดการไหลของโมเลกุลของน้ำที่วุ่นวายเพื่อให้มีความเป็นระเบียบการเปลี่ยนแปลงเอนโทรปีของน้ำ
 โครงสร้างระดับตติยภูมิของโปรตีน l l l หากชิ้นส่วนของกลุ่มที่ไม่มีขั้วของโปรตีนไม่สามารถ "ทรงกลม" ออกจากโมเลกุลได้ จะเกิดทรงกลมขึ้นซึ่งมีส่วนสำคัญ (ไม่น้อยกว่า ½) ของกลุ่มที่ไม่ชอบน้ำปรากฏขึ้นเมื่อสัมผัสกับ น้ำ สิ่งนี้เกี่ยวข้องกับการติดตั้งหน้าสัมผัสที่ไม่ชอบน้ำ หน้าสัมผัส van der Waals กองกำลัง เอนโทรปีของโปรตีนลดลงหลังจากการก่อตัวของทรงกลมมีส่วนสำคัญของความหนืดของน้ำและปฏิกิริยาที่ไม่ชอบน้ำของกลุ่มโปรตีนกับโมเลกุลของน้ำถูกทำลายความโกลาหลของส่วนที่เหลือเพิ่มขึ้นและ เอนโทรปีของน้ำเพิ่มขึ้น
โครงสร้างระดับตติยภูมิของโปรตีน l l l หากชิ้นส่วนของกลุ่มที่ไม่มีขั้วของโปรตีนไม่สามารถ "ทรงกลม" ออกจากโมเลกุลได้ จะเกิดทรงกลมขึ้นซึ่งมีส่วนสำคัญ (ไม่น้อยกว่า ½) ของกลุ่มที่ไม่ชอบน้ำปรากฏขึ้นเมื่อสัมผัสกับ น้ำ สิ่งนี้เกี่ยวข้องกับการติดตั้งหน้าสัมผัสที่ไม่ชอบน้ำ หน้าสัมผัส van der Waals กองกำลัง เอนโทรปีของโปรตีนลดลงหลังจากการก่อตัวของทรงกลมมีส่วนสำคัญของความหนืดของน้ำและปฏิกิริยาที่ไม่ชอบน้ำของกลุ่มโปรตีนกับโมเลกุลของน้ำถูกทำลายความโกลาหลของส่วนที่เหลือเพิ่มขึ้นและ เอนโทรปีของน้ำเพิ่มขึ้น

 โครงสร้างควอเทอร์นารีของโปรตีน... l l l โครงสร้างควอเทอร์นารีพบได้ในโปรตีนโอลิโกเมอร์ ซึ่งโมเลกุลประกอบด้วยหอกโพลีเปปไทด์ตั้งแต่ 2 อันขึ้นไปที่จับกันโดยไม่มีโควาเลนต์ ensemble of globules) ทำหน้าที่เป็นโมเลกุลเดี่ยว ในขณะเดียวกันผิวหนังของ globules ที่รวมกันยังคงรักษาเอกราชที่สำคัญไว้
โครงสร้างควอเทอร์นารีของโปรตีน... l l l โครงสร้างควอเทอร์นารีพบได้ในโปรตีนโอลิโกเมอร์ ซึ่งโมเลกุลประกอบด้วยหอกโพลีเปปไทด์ตั้งแต่ 2 อันขึ้นไปที่จับกันโดยไม่มีโควาเลนต์ ensemble of globules) ทำหน้าที่เป็นโมเลกุลเดี่ยว ในขณะเดียวกันผิวหนังของ globules ที่รวมกันยังคงรักษาเอกราชที่สำคัญไว้

 โครงสร้างควอเทอร์นารีของโปรตีน l หน่วยย่อยที่สร้างโครงสร้างควอเทอร์นารีของโปรตีนสามารถแตกต่างอย่างสิ้นเชิงจากการทำงานจริงของพวกมัน - โปรตีนเฮเทอโรเมอร์ริกซึ่งช่วยให้คุณรวมฟังก์ชันที่พึ่งพาซึ่งกันและกันจำนวนหนึ่งไว้ในโครงสร้างเดียว สร้างโมเลกุลที่ใช้งานได้จริงของพื้น l l แอป โปรตีนไคเนส: หน่วยย่อย C มีหน้าที่รับผิดชอบในการทำงานของเอนไซม์และหน่วยย่อย R นั้นเป็นข้อบังคับ ในโปรตีนโฮโมเมอริกหน่วยย่อยจะเหมือนกัน
โครงสร้างควอเทอร์นารีของโปรตีน l หน่วยย่อยที่สร้างโครงสร้างควอเทอร์นารีของโปรตีนสามารถแตกต่างอย่างสิ้นเชิงจากการทำงานจริงของพวกมัน - โปรตีนเฮเทอโรเมอร์ริกซึ่งช่วยให้คุณรวมฟังก์ชันที่พึ่งพาซึ่งกันและกันจำนวนหนึ่งไว้ในโครงสร้างเดียว สร้างโมเลกุลที่ใช้งานได้จริงของพื้น l l แอป โปรตีนไคเนส: หน่วยย่อย C มีหน้าที่รับผิดชอบในการทำงานของเอนไซม์และหน่วยย่อย R นั้นเป็นข้อบังคับ ในโปรตีนโฮโมเมอริกหน่วยย่อยจะเหมือนกัน
 โครงสร้างควอเทอร์นารีของโปรตีน l การสัมผัสระหว่างหน่วยย่อย - ระบบปฏิสัมพันธ์ที่ไม่ใช่โควาเลนต์ได้รับการพัฒนาแล้ว โครงสร้างควอเทอร์นารีน้อยกว่า ตติยภูมิน้อยกว่า ซึ่งหมายความว่ามีการสัมผัสที่ไม่ชอบน้ำน้อยลง
โครงสร้างควอเทอร์นารีของโปรตีน l การสัมผัสระหว่างหน่วยย่อย - ระบบปฏิสัมพันธ์ที่ไม่ใช่โควาเลนต์ได้รับการพัฒนาแล้ว โครงสร้างควอเทอร์นารีน้อยกว่า ตติยภูมิน้อยกว่า ซึ่งหมายความว่ามีการสัมผัสที่ไม่ชอบน้ำน้อยลง

 หน้าที่ของโครงสร้างหนึ่งในสี่ 1. การรวมฟังก์ชันต่างๆ ที่เกี่ยวข้องกันไว้ในโครงสร้างเดียว 2. ฟังก์ชันทางสถาปัตยกรรม 1. การสร้างโครงสร้างที่กว้างขวางให้เป็นโครงสร้างแบบพับเท่ากันเพื่อให้แน่ใจว่าโปรตีน (เฟอร์ริติน) มีสมรรถนะจำเพาะและใช้งานได้ 2. สรุปปฏิกิริยาของเอนไซม์ที่ตามมา 3. การก่อตัว ของการทำงาน
หน้าที่ของโครงสร้างหนึ่งในสี่ 1. การรวมฟังก์ชันต่างๆ ที่เกี่ยวข้องกันไว้ในโครงสร้างเดียว 2. ฟังก์ชันทางสถาปัตยกรรม 1. การสร้างโครงสร้างที่กว้างขวางให้เป็นโครงสร้างแบบพับเท่ากันเพื่อให้แน่ใจว่าโปรตีน (เฟอร์ริติน) มีสมรรถนะจำเพาะและใช้งานได้ 2. สรุปปฏิกิริยาของเอนไซม์ที่ตามมา 3. การก่อตัว ของการทำงาน
 หน้าที่ของโครงสร้างไตรมาส 3. รับประกันการมีปฏิสัมพันธ์หลายครั้งของโปรตีนด้วยโครงสร้างที่ขยายออกไป โปรตีนที่จับกับ DNA – ไดเมอร์ (?) 4. หน้าที่ด้านกฎระเบียบ การถ่ายโอนผลกระทบ (ความเสียหายต่อโครงสร้างตติยภูมิเมื่อมีการโต้ตอบกับสารตั้งต้น) จะถูกส่งจากหน่วยย่อยหนึ่งไปยังอีกหน่วยหนึ่งซึ่งนำไปสู่การฟื้นฟูโครงสร้างควอเทอร์นารีทั้งหมด
หน้าที่ของโครงสร้างไตรมาส 3. รับประกันการมีปฏิสัมพันธ์หลายครั้งของโปรตีนด้วยโครงสร้างที่ขยายออกไป โปรตีนที่จับกับ DNA – ไดเมอร์ (?) 4. หน้าที่ด้านกฎระเบียบ การถ่ายโอนผลกระทบ (ความเสียหายต่อโครงสร้างตติยภูมิเมื่อมีการโต้ตอบกับสารตั้งต้น) จะถูกส่งจากหน่วยย่อยหนึ่งไปยังอีกหน่วยหนึ่งซึ่งนำไปสู่การฟื้นฟูโครงสร้างควอเทอร์นารีทั้งหมด


 1. โปรตีนในผิวหนังแต่ละชนิดมีลักษณะเฉพาะด้วยโครงสร้างที่เป็นเอกลักษณ์ ซึ่งทำให้มั่นใจได้ถึงลักษณะเฉพาะของหน้าที่ของมัน 2 ดังนั้น การทำความเข้าใจโครงสร้างของโปรตีนต่างๆ จึงเป็นกุญแจสำคัญในการทำความเข้าใจธรรมชาติของระบบสิ่งมีชีวิต และด้วยเหตุนี้ เห็นได้ชัดว่าโดยพื้นฐานแล้ว
1. โปรตีนในผิวหนังแต่ละชนิดมีลักษณะเฉพาะด้วยโครงสร้างที่เป็นเอกลักษณ์ ซึ่งทำให้มั่นใจได้ถึงลักษณะเฉพาะของหน้าที่ของมัน 2 ดังนั้น การทำความเข้าใจโครงสร้างของโปรตีนต่างๆ จึงเป็นกุญแจสำคัญในการทำความเข้าใจธรรมชาติของระบบสิ่งมีชีวิต และด้วยเหตุนี้ เห็นได้ชัดว่าโดยพื้นฐานแล้ว
 วรรณกรรม Berezov T. T. , Korovkin B. F. เคมีชีวภาพ - อ.: แพทยศาสตร์, 2526 Metzler D. ชีวเคมี. ปฏิกิริยาเคมีในเซลล์ของสิ่งมีชีวิต ใน 3 เล่ม - T. เล่ม 1, 2. - M.: Svit, 1980 Ovchinnikov Yu. A. เคมีชีวภาพ - อ.: Prosvitnitstvo, 1987 ความรู้พื้นฐานชีวเคมี / เอ็ด. เอ.เอ. อานิซิโมวา. - M.: Vishcha Shkola, 1986 Rhys E., Strenberg M. จากเซลล์สู่อะตอม ภาพประกอบเบื้องต้นเกี่ยวกับอณูชีววิทยา - M.: Svit, 1988 Stepanov V. M. อณูชีววิทยา. โครงสร้างและหน้าที่ของโปรตีน - อ.: โรงเรียนวิชชา, พ.ศ. 2539 Pilipovich Yu. B. ความรู้พื้นฐานทางชีวเคมี - ม.: โรงเรียนวิชชา, 2536
วรรณกรรม Berezov T. T. , Korovkin B. F. เคมีชีวภาพ - อ.: แพทยศาสตร์, 2526 Metzler D. ชีวเคมี. ปฏิกิริยาเคมีในเซลล์ของสิ่งมีชีวิต ใน 3 เล่ม - T. เล่ม 1, 2. - M.: Svit, 1980 Ovchinnikov Yu. A. เคมีชีวภาพ - อ.: Prosvitnitstvo, 1987 ความรู้พื้นฐานชีวเคมี / เอ็ด. เอ.เอ. อานิซิโมวา. - M.: Vishcha Shkola, 1986 Rhys E., Strenberg M. จากเซลล์สู่อะตอม ภาพประกอบเบื้องต้นเกี่ยวกับอณูชีววิทยา - M.: Svit, 1988 Stepanov V. M. อณูชีววิทยา. โครงสร้างและหน้าที่ของโปรตีน - อ.: โรงเรียนวิชชา, พ.ศ. 2539 Pilipovich Yu. B. ความรู้พื้นฐานทางชีวเคมี - ม.: โรงเรียนวิชชา, 2536


