ساختار یک پروتئین توسط گروهی از ژن ها تعیین می شود. پروتئین ها: ساختار پروتئین اولیه، طرح ترکیب تری پپتیدی
بیلکی- ترکیبات آلی مولکولی بالا که از اسیدهای آمینه α اضافی تشکیل می شوند.
U انبار پروتئینشامل زغال سنگ، آب، نیتروژن، کیسن، سیرکا است. برخی از پروتئین ها با مولکول های دیگر مانند فسفر، آهن، روی و مس کمپلکس تشکیل می دهند.
پروتئین ها وزن مولکولی بالایی دارند: آلبومین تخم مرغ - 36000، هموگلوبین - 152000، میوزین - 500000. برای مقایسه: وزن مولکولی الکل - 46، اسید اوتیک - 60، بنزن - 78.
انبار پروتئین اسید آمینه
بیلکی- پلیمرهای غیر تناوبی مانند مونومرها اسیدهای آمینه α. 20 نوع اسید آمینه α به نام مونومرهای پروتئینی وجود دارد، اگرچه بیش از 170 نوع آن در سلول ها و بافت ها شناسایی شده است.
توجه به این نکته ضروری است که اسیدهای آمینه را می توان در بدن انسان و سایر موجودات سنتز کرد، آنها به دو دسته تقسیم می شوند: جایگزین اسیدهای آمینه- قابل سنتز اسیدهای آمینه ضروری- آنها نمی توانند سنتز کنند. آمینو اسیدهای ضروری مستقیماً از بدن در بدن یافت می شوند. گیاهان تمام انواع اسیدهای آمینه را سنتز می کنند.
ذخیره شده در انبار اسید آمینه، پروتئین ها در نوسان کامل هستند- کل مجموعه اسیدهای آمینه را غبارآلود کنید. پست تر- تمام آمینو اسیدها روزانه موجود است. از آنجایی که پروتئین ها از اسیدهای آمینه تشکیل شده اند، به آنها می گویند متاسفم. هنگامی که پروتئین ها علاوه بر اسیدهای آمینه، جزء غیر اسید آمینه دیگری (گروه مصنوعی) داشته باشند، نامیده می شوند. تاشو. گروه پروتز را می توان با فلزات (متالوپروتئین ها)، کربوهیدرات ها (گلیکوپروتئین ها)، لیپیدها (لیپوپروتئین ها)، اسیدهای نوکلئیک (نوکلئوپروتئین ها) نشان داد.
سبیل اسیدهای آمینه انتقام می گیرند: 1) گروه کربوکسیل (-COOH); 2) گروه آمینه (-NH 2). 3) رادیکال یا گروه R (مولکول رشتا). رادیکال بودوا در انواع مختلف اسیدهای آمینه متفاوت است. بسته به تعداد گروههای آمینه و گروههای کربوکسیل که وارد انبار آمینو اسید میشوند، به دو دسته تقسیم میشوند: اسیدهای آمینه خنثییک گروه کربوکسیل و یک گروه آمینه وجود دارد. اسیدهای آمینه اساسیبیش از یک گروه آمینه وجود دارد. اسیدهای آمینه اسیدیبیش از یک گروه کربوکسیل چیست؟
آمینو اسید نیمه هادی های آمفوتریکبنابراین، در غذا، بوی بد می تواند هم توسط اسیدها و هم از بازها ایجاد شود. در انواع آب، اسیدهای آمینه به اشکال مختلف یونی ظاهر می شوند.
پیوند پپتیدی
پپتیدی- ترکیبات آلی که از اسیدهای آمینه اضافی که توسط یک پیوند پپتیدی به هم متصل شده اند تشکیل می شوند.
آزاد شدن پپتیدها به دنبال واکنش تراکم اسیدهای آمینه است. هنگامی که گروه آمینه یک اسید آمینه با گروه کربوکسیل اسید آمینه دیگر برهمکنش می کند، یک پیوند کووالانسی نیتروژن-کربوهیدرات بین آنها ایجاد می شود که به نام پپتید. مهم است که مقدار اسید آمینه اضافی را که به انبار پپتید می رود جدا کنید دیپپتیدی، تریپپتیدی، تتراپپتیدیو غیره. تشکیل پیوند پپتیدی را می توان به روش های مختلفی تکرار کرد. این را به روشنی بیاور پلی پپتیدها. در یک انتهای پپتید یک گروه آمینه آزاد (به نام پایانه N) و در انتهای دیگر یک گروه کربوکسیل آزاد (به نام پایانه C) وجود دارد.

سازماندهی ساده مولکول های پروتئین
عملکردهای خاص پروتئینها به پیکربندی فضایی مولکولهای آنها بستگی دارد، علاوه بر این، سلولها با انرژی و بهطور نامرئی پروتئینها را به شکل باز که شبیه نیزه به نظر میرسد، برش میدهند، بنابراین لنجهای پلیپپتیدی تشکیل و متورم شدن ساختار و ترکیب کماهمیت مشاهده میشوند. 4 منطقه را ببینید سازماندهی وسیع پروتئین ها.
ساختار پروتئین اولیه- توالی انحلال اسیدهای آمینه اضافی از لنست پلی پپتیدی برای تشکیل یک مولکول پروتئین. پیوند بین اسیدهای آمینه پپتیدی است.

از آنجایی که یک مولکول پروتئین فقط از 10 اسید آمینه مازاد تشکیل شده است، بنابراین انواع احتمالی مولکول های پروتئینی که از نظر ترتیب افزودن اسید آمینه متفاوت هستند، 1020 است. با 20 اسید آمینه، می توانید آنها را در ترکیبات مختلف تر ترکیب کنید. حدود ده هزار پروتئین مختلف در انسان شناسایی شده است که از نوعی به نوع دیگر و همچنین از پروتئین های موجودات دیگر متفاوت است.
ساختار اولیه مولکول پروتئین نشان دهنده قدرت مولکول های پروتئین و پیکربندی وسیع آن است. جایگزینی یک اسید آمینه با دیگری در زنجیره پلی پپتیدی منجر به تغییر در قدرت و عملکرد پروتئین می شود. به عنوان مثال، جایگزینی اسید آمینه گلوتامیک در زیرواحد β هموگلوبین با والین منجر به این واقعیت می شود که مولکول هموگلوبین به طور کلی نمی تواند عملکرد اصلی خود - انتقال اسید را از دست بدهد. در چنین مواردی، افراد دچار بیماری - کم خونی سلولی داسی شکل می شوند.
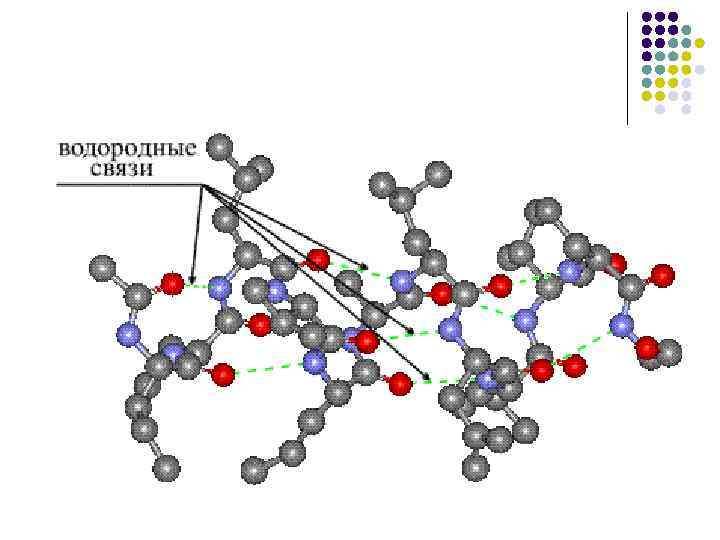
ساختار ثانویه- حنجره لنست پلی پپتیدی به صورت مارپیچی مرتب شده است (به نظر می رسد یک فنر کشیده شده است). سیم پیچ های مارپیچ توسط پیوندهای آبی تشکیل می شوند که بین گروه های کربوکسیل و گروه های آمینه در هم تنیده می شوند. تقریباً تمام گروههای CO و NH در ایجاد اتصالات آب شرکت میکنند. بوی بد نسبت به بوی پپتیدی ضعیفتر است، اما در صورت تکرار، سرشار از گاز است که به این تغییرات ماندگاری و سختی میدهد. در سطح ساختار ثانویه پروتئین ها وجود دارد: فیبروئین (بخیه، ابریشم)، کراتین (مو، ناخن)، کلاژن (تاندون).



ساختار ترتین- شلخته لانسیوگیو نیمه پتد در گلوبولی، وینیکی ونئولیکنی وینیکنی نود زویازکوف (ودنویخ، دی سولفیدنی) ایجاد میرای متقابل کمربندی توسط رادیکال های اسیدهای آمینه است. نقش اصلی ساختار ثالثی روشن توسط فعل و انفعالات آبدوست-آب گریز ایفا می شود. در مولکول های آبی، رادیکال های آبگریز تمایل به همگرایی در آب دارند و در وسط کروی گروه بندی می شوند، در حالی که رادیکال های آب دوست در نتیجه هیدراتاسیون (برهم کنش با دوقطبی های آب)، تمایل به نشستن روی سطح مولکول دارند. در برخی از پروتئین ها، ساختار سوم توسط پیوندهای کووالانسی دی سولفیدی که بین اتم های کربن دو سیستئین اضافی ایجاد می شود، تثبیت می شود. در سطح ساختار سوم آنزیم ها، آنتی بادی ها و هورمون های خاصی وجود دارد.

ساختار کواترنریو در پروتئین های تاشو، مولکول هایی که به دو یا چند گلبول تبدیل می شوند. واحدهای فرعی از طریق برهمکنش های یونی، آبگریز و الکترواستاتیک در مولکول ترکیب می شوند. گاهی اوقات وقتی ساختار چهارتایی تشکیل می شود، پیوندهای دی سولفیدی بین زیر واحدها ظاهر می شود. بزرگترین پروتئینی که یک چهارم ساختار دارد هموگلوبین. این شامل دو زیرواحد α (141 اسید آمینه مازاد) و دو زیرواحد β (146 اسید آمینه مازاد) است. یک مولکول هِم با زیرواحد پوست که مسئول رهاسازی است، مرتبط است.
اگر به هر دلیلی ترکیب پروتئین از حالت طبیعی منحرف شود، پروتئین نمی تواند عملکرد خود را حفظ کند. به عنوان مثال، علت "داستان گاو" (آنسفالوپاتی اسفنجی شکل) ترکیب غیر طبیعی پریون ها - پروتئین های سطحی سلول های عصبی است.
قدرت سنجاب ها
ذخیره اسید آمینه، ساختار مولکول پروتئین به معنای قدرت. پروتئین ها اسیدیته پایه خود را به دست می آورند، که توسط رادیکال های اسید آمینه تعیین می شود: هر چه یک پروتئین اسید آمینه اسیدی بیشتری داشته باشد، اسیدیته آن روشن تر است. ارزش دادن و آمدن H+ یعنی قدرت بافر پروتئین ها; یکی از مهم ترین بافرها هموگلوبین موجود در گلبول های قرمز است که PH خون را در سطح ثابتی حفظ می کند. - پروتئین های جداگانه (فیبرینوژن) و پروتئین های غیر جدا که به عملکردهای مکانیکی کمک می کنند (فیبرین، کراتین، کلاژن). پروتئینهای فعال به شکل شیمیایی (آنزیمها) و پروتئینهای غیرفعال شیمیایی وجود دارند که در برابر هجوم ذهنهای مختلف از دنیای خارج مقاوم و بسیار ناپایدار هستند.
عوامل خارجی (گرمایش، قرار گرفتن در معرض اشعه ماوراء بنفش، فلزات مهم و املاح آنها، تغییرات pH، تشعشع، آلودگی آب)
ممکن است باعث آسیب به سازمان ساختاری مولکول پروتئین شود. فرآیند از دست دادن ترکیب بی اهمیت، غالب یک مولکول پروتئین، نامیده می شود دناتوره سازی. دلیل دناتوره شدن پارگی رباط هایی است که ساختار پروتئین را تثبیت می کنند. ضعیفترین رباطها در کلیه پاره میشوند و وقتی ذهن قویتر باشد، قویتر پاره میشود. سپس ساختارهای چهارتایی و سپس سوم و ثانویه توسعه می یابند. تغییر پیکربندی فضایی منجر به تغییر در قدرت پروتئین می شود و در نتیجه باعث می شود پروتئین کنترل عملکردهای بیولوژیکی خود را از دست ندهد. از آنجایی که دناتوره شدن با تخریب ساختار اولیه همراه نیست، ممکن است گرگینه، هنگامی که یک خود تجدید ساختار پروتئین قدرت وجود دارد. چنین دناتوره سازی، به عنوان مثال، توسط پروتئین های گیرنده غشایی تشخیص داده می شود. فرآیند به روز رسانی ساختار پروتئین پس از دناتوراسیون نامیده می شود تجدید طبیعت. اگر تجدید پیکربندی فضایی یک پروتئین غیرممکن باشد، دناتوراسیون نامیده می شود بی اعتبار.
عملکرد پروتئین ها
| تابع | لطفا توضیحی بدهید |
|---|---|
| بودیولنا | پروتئین ها در تشکیل ساختارهای سلولی و پس از بالینی شرکت می کنند: آنها وارد غشای سلولی (لیپوپروتئین ها، گلیکوپروتئین ها)، مو (کراتین)، تاندون (کلاژن) و غیره می شوند. |
| ترانسپورنا | پروتئین خون هموگلوبین اسیدیته میافزاید و آن را از خون به تمام بافتها و اندامها منتقل میکند و از آنها دی اکسید کربن را منتقل میکند. ذخیره سازی غشای سلولی شامل پروتئین های خاصی است که انتقال فعال و کاملاً انتخابی مواد و یون های خاص از سلول به هسته بیرونی و پشت را تضمین می کند. |
| نظارتی | هورمون های پروتئینی در تنظیم فرآیندهای متابولیسم گفتار نقش دارند. به عنوان مثال، هورمون انسولین سطح گلوکز در خون را تنظیم می کند، سنتز گلیکوژن را تقویت می کند و کاهش چربی های کربوهیدرات ها را افزایش می دهد. |
| زهیسنا | هنگامی که پروتئین ها و میکروارگانیسم های خارجی (آنتی ژن ها) به بدن نفوذ می کنند، پروتئین های خاصی ایجاد می شود - آنتی بادی هایی که آنها را متصل می کنند و لایه برداری می کنند. فیبرین که با فیبرینوژن ایجاد می شود، خونریزی را کاهش می دهد. |
| دویگونا | پروتئین های کوتاه مدت اکتین و میوزین کوتاه شدن زخم ها را در حیوانات دارای سلول های غنی تضمین می کنند. |
| سیگنالنا | در غشای سطحی سلول، مولکولهای پروتئینی وجود دارد که در پاسخ به عوامل محیط خارجی، ساختار سوم خود را تغییر میدهند و در نتیجه سیگنالهایی را از محیط خارجی دریافت میکنند و دستورات را به سلول منتقل میکنند. |
| جوراب بلند کردن | در بدن حیوانات، پروتئین ها، به عنوان یک قاعده، ذخیره نمی شوند، مقصران: آلبومین تخم مرغ، کازئین شیر. با این حال، پروتئینهای موجود در بدن میتوانند با ذخیره مقدار مشخصی از گفتار تامین شوند، به عنوان مثال، هنگامی که هموگلوبین تجزیه میشود، مایع از بدن دفع نمیشود، بلکه ذخیره میشود و با پروتئین فریتین مجتمعی ایجاد میکند. |
| پر انرژی | هنگامی که 1 گرم پروتئین به محصولات نهایی تجزیه می شود، 176 کیلوژول قابل مشاهده است. در ابتدا، پروتئین ها به اسیدهای آمینه، و سپس به محصولات نهایی - آب، دی اکسید کربن و آمونیاک تجزیه می شوند. با این حال، پروتئین ها به عنوان منبع انرژی تنها در صورتی جذب می شوند که منابع دیگر (کربوهیدرات ها و چربی ها) مصرف شوند. |
| کاتالیزوری | یکی از مهمترین وظایف پروتئین هاست. حاوی پروتئین - آنزیم هایی که واکنش های بیوشیمیایی را که در سلول ها رخ می دهد سرعت می بخشد. به عنوان مثال، ریبولوز بی فسفات کربوکسیلاز تثبیت 2 را در طول فتوسنتز کاتالیز می کند. |
فرمنتی
فرمنتی، یا آنزیم ها، - دسته خاصی از پروتئین ها که کاتالیزورهای بیولوژیکی هستند. به دلیل وجود آنزیم ها، واکنش های بیوشیمیایی با سیالیت زیاد پیش می رود. سرعت واکنش های آنزیمی در ده ها هزار بار (و در میلیون ها بار) به دلیل سرعت واکنش هایی است که با مشارکت کاتالیزورهای معدنی رخ می دهد. Rechovina، همان چیزی است که آنزیم اثر خود را می دهد، نامیده می شود لایه.
آنزیم ها - پروتئین های کروی، ویژگی های بودویآنزیم ها را می توان به دو گروه ساده و پیچیده تقسیم کرد. من متاسفم تخمیرپس با ساده ترین پروتئین ها. عمدتا از اسیدهای آمینه تشکیل می شوند. آنزیم های تاشوسپس سفیده ها را تا کنید. قبل از انبار آنها علاوه بر قسمت پروتئینی، گروهی از طبیعت غیر پروتئینی نیز وجود دارد. کوفاکتور. در برخی از آنزیم ها، ویتامین ها به عنوان کوفاکتور عمل می کنند. یک مولکول آنزیم دارای بخش خاصی به نام مرکز فعال است. مرکز فعال- بخش کوچکی از آنزیم (از سه تا دوازده اسید آمینه مازاد) که سوبسترا یا سوبستراها را به کمپلکس آنزیم - سوبسترا متصل می کند. پس از اتمام واکنش، کمپلکس آنزیم-سوبسترا به آنزیم و محصول(های) واکنش تجزیه می شود. آنزیم های واقعی فعال هستند (کرم فعال) مراکز آلوستریک- بخش هایی که از تنظیم کننده ها و آنزیم های نقدینگی استفاده می شود ( آنزیم های آلوستریک).

واکنش های کاتالیز آنزیمی با موارد زیر مشخص می شود: 1) راندمان بالا، 2) گزینش پذیری دقیق و مستقیم بودن عمل، 3) اختصاصیت بستر، 4) تنظیم دقیق و دقیق. سوبسترا و ویژگی واکنش واکنش های کاتالیز آنزیمی توسط فرضیه های E. Fisher (1890) و D. Koshland (1959) توضیح داده شده است.
E. Fisher (فرضیه قفل کلید)با فرض اینکه پیکربندی گسترده مرکز فعال آنزیم و سوبسترا دقیقاً یکسان باشد. بستر همان "کلید" است، آنزیم "قفل" است.
D. Koshland (فرضیه دستی)با فرض اینکه شباهت ساختار بین سوبسترا و مرکز فعال آنزیم فقط در لحظه برهمکنش یک به یک آنها ایجاد می شود. این فرضیه نیز نامیده می شود فرضیه گونه های القا شده.
سیالیت واکنش های آنزیمی به موارد زیر بستگی دارد: 1) دما. 2) غلظت آنزیم؛ 3) غلظت بستر؛ 4) pH لازم به ذکر است که قطعات آنزیمی پروتئین هستند، پس فعالیت آنها در مغزهای طبیعی فیزیولوژیکی بالاترین است.
بیشتر آنزیم ها را فقط می توان در دماهای بین 0 تا 40 درجه سانتی گراد پردازش کرد. در این نواحی، هنگامی که دمای پوست به 10 درجه سانتیگراد می رسد، سیالیت واکنش تقریباً 2 برابر افزایش می یابد. در دمای بالاتر از 40 درجه سانتیگراد، پروتئین دچار دناتوره شده و فعالیت آنزیم کاهش می یابد. در دماهای نزدیک به نقطه انجماد، آنزیم ها غیرفعال می شوند.
با افزایش مقدار سوبسترا، سیالیت واکنش آنزیمی افزایش مییابد تا زمانی که تعداد مولکولهای زیرلایه برابر با تعداد مولکولهای آنزیم شود. با افزایش بیشتر در مقدار سوبسترا، تا زمانی که مراکز فعال آنزیم اشباع می شوند، نقدینگی افزایش نمی یابد. غلظت های بالاتر آنزیم منجر به افزایش فعالیت کاتالیزوری می شود، زیرا در یک ساعت واکنش مولکول های بیشتری در معرض سوبسترا قرار می گیرند.

برای آنزیم پوست، مقدار pH بهینه ای وجود دارد که در آن حداکثر فعالیت را نشان می دهد (پپسین - 2.0، موکوس آمیلاز - 6.8، لیپاز زیر جلدی - 9.0). در مقادیر pH بالاتر یا پایین تر، فعالیت آنزیم کاهش می یابد. در صورت تغییرات شدید pH، آنزیم دناتوره می شود.
سیالیت آنزیم های آلوستریک توسط موادی تنظیم می شود که به مراکز آلوستریک می رسند. از آنجایی که این کلمات واکنش را تسریع می کنند، به آنها بو می گویند فعال کننده ها, yakscho به galm مهار کننده ها.
طبقه بندی آنزیم ها
بر اساس نوع واکنش های شیمیایی کاتالیز شده، آنزیم ها به 6 کلاس تقسیم می شوند:
- اکسی دوکتاز(انتقال اتم ها از آب، اسید یا الکترون ها از یک ماده به ماده دیگر - دهیدروژناز)
- انتقالی(انتقال گروه های متیل، آسیل، فسفات یا آمینو از یک مولکول به مولکول دیگر - ترانس آمیناز)
- هیدرولازها(واکنش های هیدرولیز، که در آن دو محصول از بستر - آمیلاز، لیپاز) سنتز می شوند.
- لیازی(افزودن غیر هیدرولیتیک به بستر یا جدا شدن از یک گروه جدید از اتم ها، که طی آن پیوندهای C-C، C-N، C-O، C-S دکربوکسیلاز می توانند شکسته شوند)،
- ایزومراز(پربودوا داخل مولکولی - ایزومراز)،
- لیگازی(تشکیل دو مولکول در نتیجه تشکیل پیوندهای C-C، C-N، C-O، C-S - سنتتاز).
نقاشی های خود را به زیر کلاس ها و زیر کلاس ها طبقه بندی کنید. در طبقه بندی بین المللی که وجود دارد، آنزیم پوست دارای یک کد منحصر به فرد است که شامل چهار عدد است که با نقطه از هم جدا شده اند. شماره اول یک کلاس، دیگری یک زیر کلاس، سوم یک زیر کلاس، چهارم شماره سریال آنزیم در آن زیر کلاس است، مثلا کد آرژیناز 3.5.3.1 است.
رفتن به سخنرانی شماره 2"کارکردهای کربوهیدرات ها و لیپیدها چیست"
رفتن به سخنرانی شماره 4عملکرد اسیدهای نوکلئیک ATP
بیوسنتز پروتئین.
1. ساختار یک پروتئین تعیین می شود:
1) گروهی از ژن ها 2) یک ژنوم
3) یک مولکول DNA 4) کل ژن ها در یک موجود زنده
2. ژن اطلاعات مربوط به توالی مونومرها در یک مولکول را رمزگذاری می کند:
1) tRNA 2) AA 3) گلیکوژن 4) DNA
3. سه قلوها آنتی کدون نامیده می شوند:
1) DNA 2) t-RNA 3) i-RNA 4) r-RNA
4. تبادل پلاستیک به طور مهمی با واکنش ایجاد می شود:
1) از هم پاشیدگی سخنرانی های ارگانیک 2) از هم پاشیدگی سخنرانی های غیر ارگانیک
3) سنتز مواد آلی 4) سنتز مواد معدنی
5. سنتز پروتئین در سلول های پروکاریوتی رخ می دهد:
1) روی ریبوزوم ها در هسته 2) روی ریبوزوم ها در سیتوپلاسم 3) در محل سلول
4) در سطح بیرونی غشای سیتوپلاسمی
6. فرآیند پخش آغاز شده است:
1) در سیتوپلاسم 2) در هسته 3) در میتوکندری
4) روی غشای غشای آندوپلاسمی کوتاه
7. روی غشای غشای آندوپلاسمی دانه ای، سنتز رخ می دهد:
1) ATP؛ 2) در کربوهیدرات ها؛ 3) لیپیدها؛ 4) پروتئین ها
8. یک کد سه گانه:
1. یک AK 2 یک نشانه از بدن 3. آتل AK
9. سنتز پروتئین در حال حاضر کامل شده است
1. تشخیص یک کدون توسط یک آنتی کدون 2. ظهور یک "علامت تقسیم" روی ریبوزوم
3. انتقال i-RNA به ریبوزوم
10. فرآیندی که در آن اطلاعات از مولکول های DNA خوانده می شود.
1. ترجمه 2. رونویسی 3. تبدیل
11. قدرت پروتئین ها مشخص می شود...
1. ساختار ثانویه پروتئین 2. ساختار اولیه پروتئین
3. ساختار پروتئین سوم
12. فرآیندی که طی آن یک آنتی کدون به عنوان کدون در i-RNA شناخته می شود
13. مراحل بیوسنتز پروتئین.
1. رونویسی، ترجمه 2. تبدیل، ترجمه
3.تغییر سازمانی، رونویسی
14. آنتی کدون t-RNA از نوکلئوتیدهای UCG تشکیل شده است. کدام سه گانه DNA مکمل youmu است؟
1.UUG 2. TTC 3. TCG
15. تعداد t-RNA هایی که در ترجمه شرکت می کنند سنتی است:
1. کدون های i-RNA که اسیدهای آمینه را رمزگذاری می کنند. 2. مولکول های i-RNA
3 ژنی که قبل از مولکول DNA قرار دارند 4. پروتئین هایی که روی ریبوزوم ها سنتز می شوند
16. تعیین توالی نوکلئوتیدهای i-RNA در حین رونویسی از یکی از رشته های DNA: A-G-T-C-G
1) U 2) G 3) C 4) A 5) C
17. در حین تکثیر یک مولکول DNA، موارد زیر ایجاد می شود:
1) رشته ای که به قطعاتی از مولکول های دختر تقسیم شده است
2) مولکولی که از دو رشته DNA جدید تشکیل شده است
3) مولکولی که نیمی از آن از رشته های iRNA تشکیل شده است
4) یک مولکول دختر که از یک رشته قدیمی و یک رشته جدید DNA تشکیل شده است
18. الگوی سنتز یک مولکول iRNA در حین رونویسی عبارت است از:
1) کل مولکول DNA 2) سطح یکی از مولکول های DNA
3) نمودار یکی از DNA های Lancs
4) در برخی از انواع یکی از مولکول های DNA، در برخی دیگر - کل مولکول DNA.
19. فرآیند خود فرورانش یک مولکول DNA.
1. تکرار 2. تعمیر
3. تناسخ
20. در طول بیوسنتز پروتئین در سلول ها، انرژی ATP:
1) خرج کردن 2) انبار کردن
3) خرج نشده و دیده نشده
21. در سلول های جسمی یک ارگانیسم سلولی غنی:
1) مجموعه ای از ژن ها و پروتئین های مختلف 2) مجموعه ای جدید از ژن ها و پروتئین ها
3) مجموعه ای جدید از ژن ها یا مجموعه ای متفاوت از پروتئین ها
4) مجموعه ای جدید از پروتئین ها یا مجموعه ای متفاوت از ژن ها
22. یک سه گانه DNA حاوی اطلاعاتی در مورد:
1) توالی اسیدهای آمینه در یک مولکول پروتئین
2) علائم بدن 3) اسیدهای آمینه در مولکول پروتئین سنتز شده
4) ذخیره مولکول RNA
23. کدام یک از فرآیندها در سلول های هیچ عملکردی وجود ندارد:
1) سنتز پروتئین 2) متابولیسم پروتئین 3) میتوز 4) میوز
24. مفهوم "رونویسی" به این فرآیند اشاره دارد:
1) فرورانش DNA 2) سنتز i-RNA روی DNA
3) انتقال i-RNA به ریبوزوم 4) ایجاد مولکول های پروتئین بر روی پلی زوم ها
25. بخشی از یک مولکول DNA که حامل اطلاعات یک مولکول پروتئین است:
1) ژن 2) فنوتیپ 3) ژنوم 4) ژنوتیپ
26. رونویسی در یوکاریوت ها در موارد زیر رخ می دهد:
1) سیتوپلاسم 2) غشای آندوپلاسمی 3) لیزوزوم 4) هسته
27. سنتز پروتئین از:
1) شبکه آندوپلاسمی دانه ای
2) شبکه آندوپلاسمی صاف 3) هسته 4) لیزوزوم
28. یک اسید آمینه کد شده است:
1) چند نوکلئوتید 2) دو نوکلئوتید
3) یک نوکلئوتید 4) سه نوکلئوتید
29. سه گانه نوکلئوتیدهای ATC در یک مولکول DNA دارای کدون مشابه یک مولکول i-RNA است:
1) TAG 2) UAG 3) UTC 4) TsAU
30. نشانه های تقسیمکد ژنتیکی:
1. پروتئین های آهنگ را رمز می کند 2. سنتز پروتئین را تحریک می کند
3. ترویج سنتز پروتئین
31. فرآیند خود اشتراک مولکول DNA.
1. همانند سازی 2. تعمیر 3. reankornation
32. عملکرد i-RNA در فرآیند بیوسنتز.
1. ذخیره اطلاعات تراکم 2. انتقال AK به ریبوزوم
3. ارائه اطلاعات به ریبوزوم ها
33. فرآیندی که t-RNA اسیدهای آمینه را به ریبوزوم می آورد.
1. رونویسی 2. ترجمه 3. تبدیل
34. ریبوزوم ها که همان مولکول پروتئین را سنتز می کنند.
1.کروموزوم 2.پلی زوم 3.مگاکروموزوم
35. فرآیندی که طی آن اسیدهای آمینه یک مولکول پروتئین را تثبیت می کنند.
1. رونویسی 2. ترجمه 3. تبدیل
36. قبل از واکنش سنتز ماتریس می آوریم...
1. همانندسازی DNA 2. رونویسی، ترجمه 3. دو نوع صحیح هستند
37. یک سه گانه DNA حاوی اطلاعاتی در مورد:
1. توالی اسیدهای آمینه در یک مولکول پروتئین
2. محل آواز AK در لانست پروتئین
3. نشانه یک ارگانیسم خاص
4. آمینو اسیدهایی که در پروتئین لانست وجود دارند
38. Genya اطلاعات رمزگذاری شده در مورد:
1) ساختار پروتئین ها، چربی ها و کربوهیدرات ها 2) ساختار اولیه پروتئین
3) توالی نوکلئوتیدها در DNA
4) توالی اسیدهای آمینه در 2 یا بیشتر مولکول پروتئین
39. سنتز mRNA با موارد زیر آغاز می شود:
1) تقسیم DNA به دو رشته 2) برهمکنش با آنزیم RNA پلیمراز و ژن
3) فرعی ژن 4) تجزیه ژن به نوکلئوتیدها
40. رونویسی ارائه شده است:
1) در هسته 2) روی ریبوزوم ها 3) در سیتوپلاسم 4) روی کانال های EPS صاف
41. سنتز پروتئین در ریبوزوم ها در موارد زیر رخ نمی دهد:
1) سل زبودنیک 2) بژولی 3) فلای آگاریک 4) باکتریوفاژ
42. در طول ترجمه، ماتریس برای مونتاژ پروتئین پلی پپتیدی به صورت زیر است:
1) یکی از رشته های DNA 2) یکی از مولکول های DNA
3) یک مولکول iRNA 4) در برخی از رشته های DNA را تشکیل می دهد، در برخی دیگر - یک مولکول iRNA
ساختار اولیه پروتئین ها یک نیزه پلی پپتیدی خطی با آمینو اسیدها با پیوندهای پپتیدی است. ساختار اولیه ساده ترین سطح سازماندهی ساختاری یک مولکول پروتئین است. پایداری بالا توسط پیوندهای پپتیدی کووالانسی بین گروه α-آمینه یک اسید آمینه و گروه α-کربوکسیل یک اسید آمینه دیگر تضمین می شود.
اگر از نظر پیوند پپتیدی، گروه آمینه پرولین یا هیدروکسی پرولین شرکت کند، آنگاه متفاوت به نظر می رسد.

هنگامی که پیوندهای پپتیدی در کلیه تشکیل می شود، گروه کربوکسیل یک اسید آمینه فعال می شود و سپس با گروه آمینه دیگری ترکیب می شود. همچنین می توان سنتز آزمایشگاهی پلی پپتیدها را انجام داد.
پیوند پپتیدی قطعه ای از لنس پلی پپتیدی است که خود را تکرار می کند. ویژگی های کمی وجود دارد که نه تنها به شکل ساختار اولیه، بلکه به مهم ترین سطح سازمان دهی لانست پلی پپتیدی مربوط می شود:


· همسطحی - همه اتم هایی که در گروه پپتید قرار دارند در یک صفحه قرار دارند.
· اعتبار به دو شکل تشدید (کتو و انول) وجود دارد.
· جابجایی واسطه ها در اتصال صد در صد C-N.
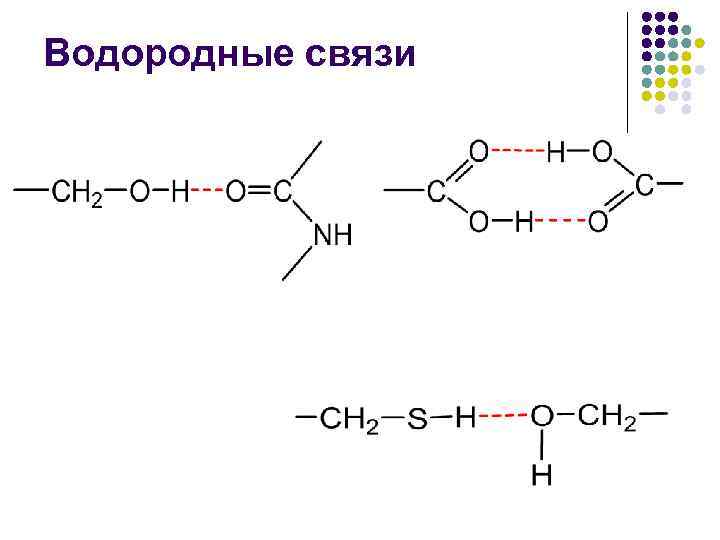
· وجود رباط های آبی که به موجب آن پوست دارای گروه های پپتیدی می تواند دو رباط آبی با گروه های دیگر از جمله رباط های پپتیدی ایجاد کند.
گروه های پپتیدی با گروه آمینه پرولین یا هیدروکسی پرولین مرتبط هستند. بوی تعفن ساختمان فقط با یک صدای آبکی (چیز شگفت انگیز) ایجاد می شود. این در ساختار ثانویه قالب گیری شده پروتئین نشان داده شده است. ترکیب پلی پپتیدی که حاوی پرولین یا هیدروکسی پرولین است، به راحتی از بین می رود و مانند همیشه با سایر پیوندهای آبی قابل حذف نیست.
طرح فرمولاسیون تری پپتید:

سطوح سازماندهی فضایی پروتئین ها: ساختار ثانویه پروتئین ها: مفاهیم مارپیچ α و توپ تا شده β. ساختار سوم پروتئین ها: مفاهیمی در مورد پروتئین بومی و دناتوره شدن پروتئین ساختار چهارتایی پروتئین ها شبیه هموگلوبین است.
ساختار ثانویه پروتئینتحت ساختار ثانویه پروتئین، ما روشی را درک می کنیم که در آن لنس پلی پپتیدی در یک ساختار مرتب مرتب شده است. با توجه به پیکربندی، عناصر زیر از ساختار ثانویه قابل مشاهده هستند: α -مارپیچ β توپ قطعات انبار.
مدل بودوی مارپیچ α، که از تمام قدرت پیوند پپتیدی محافظت می کند، توسط L. Pauling و R. Corey (1949 - 1951) شکسته شد.
به کودک 3، آنمودار نشان داده شده است α - مارپیچی، که اطلاعاتی در مورد پارامترهای اصلی لانست پلی پپتیدی میسوزاند α - یک مارپیچ به گونه ای که چرخش های مارپیچ منظم باشد، بنابراین پیکربندی مارپیچ دارای تقارن پیچ است (شکل 3، ب). روی یک چرخش چرمی α مارپیچ ها 3.6 اسید آمینه مازاد دارند. بین پیچ ها بایستید یا لبه مارپیچ 0.54 نانومتر می شود، جایی که چرخش بعدی به 26 درجه می رسد. شکل دادن و پیرایش α - پیکربندی مارپیچی توسط ساختار رباط های آبی که بین گروه های پپتیدی پوست ایجاد می شود تشکیل می شود. n- برو تا ( پ+ 3) اسید آمینه مازاد. اگرچه انرژی رباط های آب کم است، اما مقدار آنها برای ایجاد یک اثر انرژی قابل توجه است، در نتیجه α - پیکربندی مارپیچ برای تکمیل پایه. رادیکال های بیولوژیکی اسیدهای آمینه اضافی در این ماده شرکت نمی کنند α -پیکربندی مارپیچی، بنابراین تمام اسیدهای آمینه مازاد در آن وجود دارد α -مارپیچ ها برابرند.
پروتئین های طبیعی کمتر راست دست هستند α - مارپیچ.
توپ تا شده β- یکی دیگر از عناصر ساختار ثانویه. در صفحه مدیریت α -مارپیچ β -توپ تا شده شکل خطی دارد نه شکل strizhnevuyu (شکل 4). ساختار خطی با منشاء پیوندهای آبی بین گروه های پپتیدی که در بخش های مختلف لنس پلی پپتیدی قرار دارند تعیین می شود. این نمودارها به نظر می رسد نزدیک به سطح اتصال آب بین - C = O و HN - گروه (0.272 نانومتر).

کم اهمیت 4. تصویر شماتیک β
- توپ مکرر انبار (فلش ها نشان می دهد
 در مورد لانست پلی پپتیدی)
در مورد لانست پلی پپتیدی)
کم اهمیت 3. طرح ( آ) آن مدل ( ب) α -مارپیچ
ساختار ثانویه پروتئین به عنوان ساختار اولیه در نظر گرفته می شود. اسید آمینه اضافی در مقادیر مختلف قبل از تشکیل رباطهای آبی ایجاد میشود که سپس در مخلوط ریخته میشود. α -مارپیچ یا β -شارو. اسیدهای آمینه محلول در مارپیچ عبارتند از آلانین، گلوتامیک اسید، گلوتامین، لوسین، لیزین، متیونین و هیستیدین. اگر یک قطعه پروتئین در درجه اول از اسیدهای آمینه اضافی تشکیل شود، در این مرحله تشکیل می شود α -مارپیچ والین، ایزولوسین، ترئونین، تیروزین و فنیل آلانین محلول را فراهم می کنند. β - توپ های لانست پلی پپتیدی. ساختارهای نامرتب بر روی پلات های لنس پلی پپتیدی ظاهر می شوند، جایی که بقایای اسیدهای آمینه مانند گلیسین، سرین، اسید آسپارتیک، آسپاراژین، پرولین متمرکز شده اند.
در پروتئین های غنی، بلافاصله α -مارپیچ، که β -شری. بخشی از پیکربندی مارپیچ در پروتئین های مختلف متفاوت است. بنابراین، پروتئین گوشت پارامیوزین عملاً 100٪ مارپیچی است. نسبت بالایی از پیکربندی مارپیچ در میوگلوبین و هموگلوبین (75٪). با این حال، در تریپسین و ریبونوکلئاز، بخش قابل توجهی از لانژوگ پلی پپتیدی در شاروات قرار می گیرد. β -سازه های. پروتئین های بافت های پشتیبان - کراتین (پروتئین های مو)، کلاژن (پروتئین های پوست و تاندون) - متورم می شوند. β -پیکربندی لنج های پلی پپتیدی
ساختار پروتئین سومساختار سوم پروتئین راهی برای قرار دادن لنس پلی پپتیدی در فضا است. برای اینکه پروتئین قدرتی به قدرت عملکردی خود بیافزاید، لانست پلی پپتیدی موظف است به تنهایی در فضا بسوزد و یک ساختار از نظر عملکردی فعال تشکیل دهد. این ساختار نامیده می شود بومی صرف نظر از تعداد زیادی از ساختارهای فضایی که از نظر تئوری برای لانست پلی پپتیدی اطراف ممکن است، پروتئین حنجره باید به یک پیکربندی بومی منفرد تبدیل شود.
ساختار سوم پروتئین را با فعل و انفعالاتی که بین رادیکال های بیولوژیکی اسیدهای آمینه اضافی بخش های مختلف لنس پلی پپتیدی رخ می دهد، تثبیت کنید. این تعاملات را می توان به نقاط قوت و ضعف تقسیم کرد.
فعل و انفعالات قوی شامل پیوندهای کووالانسی بین اتم های گوگرد سیستئین اضافی است که در قسمت های مختلف لانس پلی پپتیدی قرار دارند. در غیر این صورت، چنین پیوندهایی را پل های دی سولفیدی می نامند. شکل گیری یک پل دی سولفیدی را می توان به صورت زیر نشان داد:
علاوه بر پیوندهای کووالانسی، ساختار سوم مولکول پروتئین تحت تأثیر برهمکنشهای ضعیف است که بر اساس ماهیت خود به قطبی و غیرقطبی تقسیم میشوند.
فعل و انفعالات قطبی شامل اتصالات یونی و آب است. فعل و انفعالات یونی بر اثر تماس بین گروه های با بار مثبت از رادیکال های زائد لیزین، آرژنین، هیستیدین و گروه های COOH با بار منفی اسیدهای آسپارتیک و گلوتامیک ایجاد می شود. اتصالات آب بین گروه های عملکردی رادیکال های بیولوژیکی اسید آمینه اضافی رخ می دهد.
فعل و انفعالات غیرقطبی یا واندروالسی بین رادیکال های کربوهیدرات باقی مانده اسیدهای آمینه مانع از تشکیل هسته آبگریز (لکه های چربی) در وسط گلبول پروتئین، زیرا رادیکال های کربوهیدرات از تماس با آب ناپدید می شوند. هر چه یک پروتئین حاوی اسیدهای آمینه غیرقطبی بیشتری باشد، نقش پیوندهای واندروالسی در ساختار سوم شکل آن بیشتر است.
اتصالات عددی بین رادیکال های پروتئینی اسیدهای آمینه اضافی، پیکربندی فضایی مولکول پروتئین را نشان می دهد (شکل 5).

کم اهمیت 5. انواع رباط هایی که از ساختار سوم پروتئین حمایت می کنند:
آ- مخلوط دی سولفید؛ ب -پیوند یونی؛ ج، د -اتصالات آب؛
د -اوراق قرضه واندروالس
ساختار سوم پروتئین تازه گرفته شده منحصر به فرد است، همانطور که ساختار اولیه منحصر به فرد است. فقط فضای مناسب برای قرار دادن پروتئین آن را فعال نگه می دارد. تخریب گسترده ساختار سوم منجر به تغییر در سطح پروتئین و از دست دادن فعالیت بیولوژیکی می شود.
ساختار چهارتایی پروتئینپروتئین هایی با وزن مولکولی بیش از 100 کیلو دالتون معمولاً از چندین لنج پلی پپتیدی با وزن مولکولی کوچک تشکیل شده اند. ساختاری که از تعداد زیادی لنج پلی پپتیدی تشکیل شده است که موقعیت کاملاً ثابتی را اشغال می کنند ، ظاهراً یکی از دیگری ، در نتیجه پروتئین فعالیت متفاوتی دارد ، ساختار چهارتایی پروتئین نامیده می شود. پروتئینی که ساختار یک چهارم دارد نامیده می شود اپی مولکولیا چیز دیگر مولتیمر ، و انبارهای لنتس پلی پپتیدی آن به وضوح مشخص است زیر واحدها یا چیز دیگر پروتومیرها . قدرت مشخصه پروتئین هایی با ساختار چهارتایی آنهایی هستند که زیر واحد آنها فعالیت بیولوژیکی ندارد.
تثبیت ساختار چهارتایی پروتئین از طریق برهمکنشهای قطبی بین رادیکالهای اسید آمینه که در سطح زیرواحدها قرار دارند به دست میآید. چنین تعاملاتی برای کاهش زیرواحدهای یک مجموعه سازمان یافته مهم هستند. به تقسیمات زیر واحدهایی که برهم کنش دارند، میدان تماسی می گویند.
نمونه کلاسیک پروتئینی که ساختار یک چهارم دارد هموگلوبین است. مولکول هموگلوبین با وزن مولکولی 68000. این مولکول از چهار زیر واحد از دو نوع مختلف تشکیل شده است. α і β / α -این زیر واحد از 141 اسید آمینه تشکیل شده است، a β - IZ 146. ساختار سوم α - І β - زیرواحد مشابه وزن مولکولی آن است (17000 So). زیرواحد پوست برای انتقام از گروه پروتز - هم . قطعات هم در پروتئین های دیگر (سیتوکروم، میوگلوبین) نیز وجود دارند که بیشتر تشکیل می شوند، اما ما می خواهیم به طور خلاصه ساختار آن ها را مورد بحث قرار دهیم (شکل 6). هم گروهی یک سیستم حلقوی همسطح چین خورده است که از یک اتم مرکزی تشکیل شده است که پیوندهای هماهنگی با مقداری پلیمر اضافی ایجاد می کند که توسط سایت های متان (=CH -) به هم متصل شده اند. در هموگلوبین، سطح اکسیداسیون شروع به رخ دادن می کند (2+).
چندین زیر واحد - دو α و دو β - در یک ساختار واحد متحد شوند به گونه ای که α زیرواحدها فقط تماس دارند β -زیر واحدها و به طور کلی (شکل 7).

کم اهمیت 6. ساختار هموگلوبین هِم

کم اهمیت 7. نمایش شماتیک ساختار چهارتایی هموگلوبین:
آهن - هموگلوبین هم
همانطور که از نوزاد 7 مشاهده می شود، یک مولکول هموگلوبین قادر به انتقال 4 مولکول اسید است. هم اتصال و هم آزاد شدن اسیدیته با تغییرات ساختاری در ساختار همراه است α - І β زیرواحد هموگلوبین و انحلال متقابل آنها در اپی مولکول این واقعیت تأیید می کند که ساختار چهارم پروتئین دیگر کاملاً سفت نیست.
اطلاعات مشابه
یکی از ویژگی های پروتئین ها سازماندهی ساختاری پیچیده آنهاست. همه پروتئین ها دارای ساختار اولیه، ثانویه و سوم هستند و آنهایی که دارای دو یا چند PPC هستند ساختار چهارتایی (QS) دارند.
ساختار پروتئین اولیه (PSB) – این ترتیب رسوب (توالی) اسیدهای آمینه اضافی در PPC است.
اما به دلیل فراوانی و ترکیب اسید آمینه، پروتئین ها می توانند انواع مختلفی داشته باشند. به عنوان مثال، از دو اسید آمینه می توانید 2 دی پپتید مختلف را ترکیب کنید:
با تعداد اسیدهای آمینه بیش از 20، تعداد ترکیبات ممکن کمتر از 210 18 است. و اگر می دانید که در PPC اسید آمینه پوست می تواند بیش از یک بار حساس شود، مهم است که از تعداد گزینه های ممکن محافظت کنید.
اهمیت ساختار پروتئین اولیه (PPS).
PSB پروتئین ها را می توان برای کمک محاسبه کرد فنیل تیوهیدانتوئین
روش
. این روش مبتنی بر واکنش متقابل است فنیل ایزوتیوسیانات
(FITC) با α-AA. در نتیجه مجموعه ای از دو رابطه ایجاد می شود FITC-AK
.
به عنوان مثال، بیایید به پپتید نگاه کنیم ![]() با استفاده از این روش، از PSB برای تعیین توالی اسیدهای آمینه اضافی استفاده می شود.
با استفاده از این روش، از PSB برای تعیین توالی اسیدهای آمینه اضافی استفاده می شود.
FITC با اسید آمینه پایانی (a) برهمکنش می کند. مجتمع در حال تاسیس است FTG-aکه به مخلوط اضافه می شود و نشان دهنده قدرت اسید آمینه است. آ. به عنوان مثال، tse - asn
![]() و غیره. تمام اسیدهای آمینه دیگر را به طور مداوم تقویت و شناسایی کنید. این یک فرآیند پر زحمت است. یک پروتئین PSB با اندازه متوسط چندین ماه دوام می آورد.
و غیره. تمام اسیدهای آمینه دیگر را به طور مداوم تقویت و شناسایی کنید. این یک فرآیند پر زحمت است. یک پروتئین PSB با اندازه متوسط چندین ماه دوام می آورد.
رمزگشایی PSB دارای اولویت است سنگرو(1953)، که کشف انسولین PSB (برنده جایزه نوبل) است. مولکول انسولین از 2 PPC - A و B تشکیل شده است.

A-Lancet از 21 اسید آمینه تشکیل شده است، لانست - از 30. PPC توسط سایت های دی سولفید به یکدیگر متصل می شود. تعداد پروتئین هایی که PSB های آنها امروزه شناسایی شده است به 1500 می رسد. تغییرات جزئی در ساختار اولیه می تواند به طور قابل توجهی قدرت پروتئین را تغییر دهد. در گلبول های قرمز افراد سالم، HbA وجود دارد - در صورت جایگزینی در -lancer HbA، در موقعیت ششم. gluبر شفتسرزنش بیماری جدی کم خونی داسی شکل، برای هر کودکی که با این ناهنجاری متولد شود، زود می میرد. از سوی دیگر، گزینه های احتمالی برای تغییر PSB وجود دارد که توسط هیچ مرجع فیزیکوشیمیایی و بیولوژیکی مشخص نشده است. مثلا، HbC به جای گلولیز در جایگاه ششم b-lance قرار دارد، HbC ممکن است با قدرت آن بر HbA رقابت نکند و افرادی که چنین Hb را در گلبول های قرمز خود حمل می کنند عملا سالم هستند.
پایداری PSBعمدتاً توسط پیوندهای پپتیدی کووالانسی داخلی و به عبارت دیگر با پیوندهای دی سولفیدی تأمین می شود.
ساختار ثانویه پروتئین (PSB).
PPV پروتئین ها انعطاف پذیری زیادی از خود نشان می دهد و یک ساختار جادار مشخص ایجاد می کند ساختار. در پروتئین ها، 2 سطح از چنین ترکیبی وجود دارد - این VSB و ساختار سوم است. (TBB).
VSB – این پیکربندی PPC، نحوه قرار دادن یا پیچاندن آن به هر ترکیبی، با برنامه موجود در P سازگار است. SB.
سه نوع اصلی VSB وجود دارد:
1) -مارپیچ;
2) ب-ساختار(انواع قطعات توپ یا ورق قطعات را تا کنید)؛
3) توپ بدون لک
-مارپیچ .
این مدل توسط V. Pauling پیشنهاد شد. این برای پروتئین های کروی بهترین است. برای هر سیستمی، پایدارترین حالت، حالتی است که حداقل انرژی آزاد را نشان دهد. برای پپتیدها، این وضعیت زمانی رخ میدهد که گروههای CO و NH توسط یک پیوند آبی ضعیف به یکدیگر متصل شوند. U آ -مارپیچ گروه NH از اولین باقی مانده اسید آمینه با گروه CO از اسید آمینه چهارم تعامل دارد. در نتیجه، ستون فقرات پپتیدی یک مارپیچ ایجاد می کند که 3.6 AA اضافی روی آن روی پوست می افتد.
1 مارپیچ مارپیچی (1 دور) = 3.6 AC = 0.54 نانومتر، برش - 26 درجه
چرخش PPC در پشت فلش سال مشاهده می شود، به طوری که مارپیچ به سمت راست حرکت می کند. از طریق پوسته 5 دور (18 AC؛ 2.7 نانومتر) پیکربندی PPC تکرار می شود.
تثبیت کننده VSBاولاً قبل از پیوندهای آبی و در دیگری - پپتید و دی سولفید. پیوندهای آبی 10-100 برابر ضعیف تر از پیوندهای شیمیایی استاندارد هستند. با این حال، مقدار زیاد بوی بد آنها سختی و فشرده بودن VSB را تضمین می کند. L-Lancelet ها به صورت مارپیچی هستند که تا حد زنگ زدن اریب هستند و در امتداد اضلاع و محورهای مختلف پخش می شوند.
ب -ساختار .
اینها قسمت های تا شده طرح PPC هستند که شبیه به شکل یک برگ است که مانند آکاردئون تا شده است. توپ های PPC می توانند موازی باشند، زیرا حمله از انتهای N یا C شروع می شود.
از آنجایی که لنزهای باریک توپ با انتهای نزدیک N-C و C-N جهت گیری شده است، پس بو نامیده می شود. ضد موازی.

 موازی
موازی
![]() ضد موازی
ضد موازی
تشکیل پیوندهای آبی، مانند مارپیچ a، بین گروههای CO و NH اتفاق میافتد.

 مطالعه سازماندهی ساختاری پروتئینها یکی از مشکلات اصلی بیوشیمی مدرن است که اهمیت علمی و عملی مهمی برای درک گسترش زیاد عملکرد پروتئینها دارد. آیا در یک مولکول پروتئین ده ها یا صدها اسید آمینه وجود دارد؟
مطالعه سازماندهی ساختاری پروتئینها یکی از مشکلات اصلی بیوشیمی مدرن است که اهمیت علمی و عملی مهمی برای درک گسترش زیاد عملکرد پروتئینها دارد. آیا در یک مولکول پروتئین ده ها یا صدها اسید آمینه وجود دارد؟
 پروتئین های امیل فیشر پلی پپتیدهای چین خورده ای هستند که در آن اسیدهای آمینه توسط پیوندهای پپتیدی (R-CO-NH-R) به یکدیگر متصل می شوند که در طی برهمکنش گروه های کربوکسیل و آمینو اسیدهای آمینه تشکیل می شوند.
پروتئین های امیل فیشر پلی پپتیدهای چین خورده ای هستند که در آن اسیدهای آمینه توسط پیوندهای پپتیدی (R-CO-NH-R) به یکدیگر متصل می شوند که در طی برهمکنش گروه های کربوکسیل و آمینو اسیدهای آمینه تشکیل می شوند.
 اثبات تجربی نظریه l l l پروتئین های طبیعی دارای تعداد کمی گروه تیتر شده – COOH و –NH 2 هستند. در طی فرآیند هیدرولیز پروتئین، تعداد استوکیومتری از گروه های تیتر شده – COOH و –NH 2 تحت تأثیر پروتئولیت ها ایجاد می شود. با وجود آنزیم های خاص، پروتئین ها به قطعات کاملاً مشخص (پلی پپتیدها) شکسته می شوند. تجزیه و تحلیل اشعه ایکس ساختار پلی پپتیدی را تأیید می کند. پروتئین تأیید اصلی نظریه پلی پپتیدی پروتئین، امکان سنتز پلی پپتیدها و پروتئین ها از قبلاً شناخته شده است. پروتئین (انسولین، لیزوزیم، ریبونوکلئوس) با روش های شیمیایی.
اثبات تجربی نظریه l l l پروتئین های طبیعی دارای تعداد کمی گروه تیتر شده – COOH و –NH 2 هستند. در طی فرآیند هیدرولیز پروتئین، تعداد استوکیومتری از گروه های تیتر شده – COOH و –NH 2 تحت تأثیر پروتئولیت ها ایجاد می شود. با وجود آنزیم های خاص، پروتئین ها به قطعات کاملاً مشخص (پلی پپتیدها) شکسته می شوند. تجزیه و تحلیل اشعه ایکس ساختار پلی پپتیدی را تأیید می کند. پروتئین تأیید اصلی نظریه پلی پپتیدی پروتئین، امکان سنتز پلی پپتیدها و پروتئین ها از قبلاً شناخته شده است. پروتئین (انسولین، لیزوزیم، ریبونوکلئوس) با روش های شیمیایی.

 ویژگی های ساختاری لنس پپتیدی پیوند پپتید تقریباً 10٪ کوتاهتر از پیوند -C-N- است و ویژگی یک پیوند "مکرراً غوطه ور" -C=N- توسط L. Pauling و R. Corey را دارد. توسعه یافته در 1948-1955 صفحات. ماهیت خاص پیوند C-N را با "رزونانس" بین دو شکل توضیح دهید
ویژگی های ساختاری لنس پپتیدی پیوند پپتید تقریباً 10٪ کوتاهتر از پیوند -C-N- است و ویژگی یک پیوند "مکرراً غوطه ور" -C=N- توسط L. Pauling و R. Corey را دارد. توسعه یافته در 1948-1955 صفحات. ماهیت خاص پیوند C-N را با "رزونانس" بین دو شکل توضیح دهید
 ویژگیهای ساختاری پپتید لنس l l l به عبارت دیگر، در پروتئینها و پپتیدها، پیوندهای C-N اغلب از طریق برهمکنش جفت الکترونهای اتم نیتروژن با سیستم الکترونیکی گروه کربونیل متعدد هستند، که منجر به مشکل میشود. تشکیل tannya navkolo zvjazku C–N Zazvichay پپتید zv' پیکربندی ترانس با 2.6 کیلوکالری در مول (10.878 کیلوژول بر مول) نسبت به پیکربندی cis بارزتر است، زیرا نزدیکی اتم های کربن در پیکربندی cis کار را دشوار می کند.
ویژگیهای ساختاری پپتید لنس l l l به عبارت دیگر، در پروتئینها و پپتیدها، پیوندهای C-N اغلب از طریق برهمکنش جفت الکترونهای اتم نیتروژن با سیستم الکترونیکی گروه کربونیل متعدد هستند، که منجر به مشکل میشود. تشکیل tannya navkolo zvjazku C–N Zazvichay پپتید zv' پیکربندی ترانس با 2.6 کیلوکالری در مول (10.878 کیلوژول بر مول) نسبت به پیکربندی cis بارزتر است، زیرا نزدیکی اتم های کربن در پیکربندی cis کار را دشوار می کند.
 ویژگی های ساختاری لنس پپتید چرخش ها حول دو پیوند ساده (N-C و C-C 1) امکان پذیر است که به اتم نامتقارن = 180 - تبدیل می شوند.
ویژگی های ساختاری لنس پپتید چرخش ها حول دو پیوند ساده (N-C و C-C 1) امکان پذیر است که به اتم نامتقارن = 180 - تبدیل می شوند.
 ویژگی های ساختاری لانست پپتیدی l l به دلیل تعامل بین مدافعان در لانست پپتیدی، آنها نمی توانند معنایی پیدا کنند - فقط مناطق گسسته خاصی برای آنها مجاز است که نشان دهنده قدرت انرژی است برخی از ترکیبات پپتید lanjug Kuti دارای روابط متقابل هستند. تغییر یکی از آنها، تغییر دیگری دشوارتر است
ویژگی های ساختاری لانست پپتیدی l l به دلیل تعامل بین مدافعان در لانست پپتیدی، آنها نمی توانند معنایی پیدا کنند - فقط مناطق گسسته خاصی برای آنها مجاز است که نشان دهنده قدرت انرژی است برخی از ترکیبات پپتید lanjug Kuti دارای روابط متقابل هستند. تغییر یکی از آنها، تغییر دیگری دشوارتر است

 l K. W. Linderström-Lang 4 سطح سازماندهی مولکول های پروتئین را متمایز کرد - ساختارهای اولیه، ثانویه، سوم و چهارم. اگرچه این دسته بندی ها منسوخ شده اند، اما فعلاً استفاده از آنها ادامه خواهد داشت.
l K. W. Linderström-Lang 4 سطح سازماندهی مولکول های پروتئین را متمایز کرد - ساختارهای اولیه، ثانویه، سوم و چهارم. اگرچه این دسته بندی ها منسوخ شده اند، اما فعلاً استفاده از آنها ادامه خواهد داشت.

 ساختار اولیه یک پروتئین l l دنباله ای از بقایای اسید آمینه در یک لانست پلی پپتیدی توسط ژن ساختاری این پروتئین رمزگذاری شده است و حاوی هر چیزی است که برای خودسازماندهی ساختار وسیع آن لازم است همه پروتئین ها برای خود با این ساختار اولیه رقابت می کنند. تعداد بالقوه چنین ساختارهایی به تعداد انواع مختلف پروتئین در همه موجودات زنده محدود نمی شود.حدود 1010-1012
ساختار اولیه یک پروتئین l l دنباله ای از بقایای اسید آمینه در یک لانست پلی پپتیدی توسط ژن ساختاری این پروتئین رمزگذاری شده است و حاوی هر چیزی است که برای خودسازماندهی ساختار وسیع آن لازم است همه پروتئین ها برای خود با این ساختار اولیه رقابت می کنند. تعداد بالقوه چنین ساختارهایی به تعداد انواع مختلف پروتئین در همه موجودات زنده محدود نمی شود.حدود 1010-1012
 درک عملکرد بیولوژیکی و مکانیسم مولکولی فعالیت فیزیولوژیکی یک پروتئین بدون آگاهی دقیق از وجود آن غیرممکن است. بررسی ساختار اولیه پروتئینهای «جهشزا» درک ماهیت بیماریهای رکودی را در سطح مولکولی ممکن میسازد.
درک عملکرد بیولوژیکی و مکانیسم مولکولی فعالیت فیزیولوژیکی یک پروتئین بدون آگاهی دقیق از وجود آن غیرممکن است. بررسی ساختار اولیه پروتئینهای «جهشزا» درک ماهیت بیماریهای رکودی را در سطح مولکولی ممکن میسازد.
 روشهای تعیین ساختار اولیه l آماده سازی پروتئین قبل از تجزیه و تحلیل ساختار اولیه با هدف به حداقل رساندن تزریق سایر سطوح بالاتر سازمان آن انجام می شود به عبارت دیگر ممکن است موضوع آنالیز بدون هیچ گونه لنس این پروتئین دچار اختلال شود. پیوندهای کووالانسی عرضی (به عنوان مثال دی سولفید) به طوری که همه پیوندهای پپتیدی هم برای معرف های شیمیایی و هم برای آنزیم ها در دسترس هستند.
روشهای تعیین ساختار اولیه l آماده سازی پروتئین قبل از تجزیه و تحلیل ساختار اولیه با هدف به حداقل رساندن تزریق سایر سطوح بالاتر سازمان آن انجام می شود به عبارت دیگر ممکن است موضوع آنالیز بدون هیچ گونه لنس این پروتئین دچار اختلال شود. پیوندهای کووالانسی عرضی (به عنوان مثال دی سولفید) به طوری که همه پیوندهای پپتیدی هم برای معرف های شیمیایی و هم برای آنزیم ها در دسترس هستند.
 l l پروتئین ابتدا مسئول تمام دناتوره شدن عمیق و از دست دادن ساختارهای چهارم، سوم و در صورت امکان ثانویه است.
l l پروتئین ابتدا مسئول تمام دناتوره شدن عمیق و از دست دادن ساختارهای چهارم، سوم و در صورت امکان ثانویه است.
 l برای جلوگیری از تشکیل پیوندهای دی سولفیدی، گروه های سولفیدریل را دوباره با اسید یدوکتیک اضافی مسدود کنید: R-SH + I-CH 2-COOH R-S-CH 2-COOH + HI
l برای جلوگیری از تشکیل پیوندهای دی سولفیدی، گروه های سولفیدریل را دوباره با اسید یدوکتیک اضافی مسدود کنید: R-SH + I-CH 2-COOH R-S-CH 2-COOH + HI
 سپس، هیدرولیز انتخابی پروتئین دناتوره شده برای حذف سیستم پپتیدی که همپوشانی دارند، انجام می شود. هیدرولیز با آنزیم ها انجام می شود: l l تریپسین (اتصالات ایجاد شده توسط گروه های کربوکسیل اسیدهای آمینه معطر)، کیموتریپسین (اتصالات ایجاد شده توسط گروه های -COOH اسیدهای آمینه معطر) و غیره. معرف های شیمیایی: بروموسیان Br - C N (بایدکننده ها، ترکیب شده با گروه ها - COOH met) و غیره.
سپس، هیدرولیز انتخابی پروتئین دناتوره شده برای حذف سیستم پپتیدی که همپوشانی دارند، انجام می شود. هیدرولیز با آنزیم ها انجام می شود: l l تریپسین (اتصالات ایجاد شده توسط گروه های کربوکسیل اسیدهای آمینه معطر)، کیموتریپسین (اتصالات ایجاد شده توسط گروه های -COOH اسیدهای آمینه معطر) و غیره. معرف های شیمیایی: بروموسیان Br - C N (بایدکننده ها، ترکیب شده با گروه ها - COOH met) و غیره.
 مرحله اول - تفکیک پپتیدهای جدا شده - با روش های کروماتوگرافی انجام می شود، مرحله باقی مانده - تعیین ساختار اولیه l با اهمیت وزن مولکولی، ترکیب اسید آمینه، اسید آمینه N- و اضافی اسید آمینه پایانی آغاز می شود. عامل دیگری را انتخاب کنید و نوع دیگری و متفاوت از اولین مجموعه قطعات پپتیدی را انتخاب کنید که برای تعیین ترتیب توزیع اسید آمینه در پروتئین تقسیم و تجزیه و تحلیل می شود.
مرحله اول - تفکیک پپتیدهای جدا شده - با روش های کروماتوگرافی انجام می شود، مرحله باقی مانده - تعیین ساختار اولیه l با اهمیت وزن مولکولی، ترکیب اسید آمینه، اسید آمینه N- و اضافی اسید آمینه پایانی آغاز می شود. عامل دیگری را انتخاب کنید و نوع دیگری و متفاوت از اولین مجموعه قطعات پپتیدی را انتخاب کنید که برای تعیین ترتیب توزیع اسید آمینه در پروتئین تقسیم و تجزیه و تحلیل می شود.

 روش F. Sanger l U 1945 r. بیوشیمیدان انگلیسی F. Sanger یکی از اولین روشها را برای شناسایی اسیدهای آمینه N ترمینال ایجاد کرد
روش F. Sanger l U 1945 r. بیوشیمیدان انگلیسی F. Sanger یکی از اولین روشها را برای شناسایی اسیدهای آمینه N ترمینال ایجاد کرد
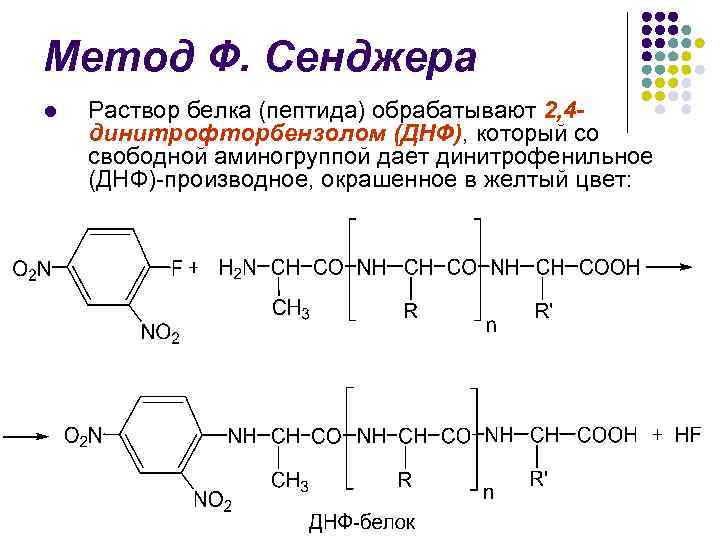 F. روش سانگر l پروتئین (پپتید) با 2، 4 دینیتروفلوئوروبنزن (DNF) تیمار میشود که از گروه آمینه آزاد دینیتروفنیل (DNF) حاوی رنگ زرد میدهد:
F. روش سانگر l پروتئین (پپتید) با 2، 4 دینیتروفلوئوروبنزن (DNF) تیمار میشود که از گروه آمینه آزاد دینیتروفنیل (DNF) حاوی رنگ زرد میدهد:
 روش F. Sanger l l هیدرولیز اسید تهاجمی (5.7 N HCl) منجر به تخریب پیوندهای پپتیدی و ایجاد اسید آمینه N ترمینال DNP مانند می شود.
روش F. Sanger l l هیدرولیز اسید تهاجمی (5.7 N HCl) منجر به تخریب پیوندهای پپتیدی و ایجاد اسید آمینه N ترمینال DNP مانند می شود.
 روش Dansilny l Rozrobleny در سال 1963. بیوشیمیدان انگلیسی W. Gray و B. Hartley
روش Dansilny l Rozrobleny در سال 1963. بیوشیمیدان انگلیسی W. Gray و B. Hartley
 روش دانسیل l مرحله اول - واکنش دانسیل کلرید (1-دی متیل آمینوافتالین-5-سولفوکلرید) با یک پپتید (یا پروتئین) گروه آمینه بدون پروتون با یک پپتید دانسیل اصلاح شده (پپتید DNS)
روش دانسیل l مرحله اول - واکنش دانسیل کلرید (1-دی متیل آمینوافتالین-5-سولفوکلرید) با یک پپتید (یا پروتئین) گروه آمینه بدون پروتون با یک پپتید دانسیل اصلاح شده (پپتید DNS)

 قویترین روش حذف پروتئینهای DNS تحت هیدرولیز در حضور 5، 7 n است. HCl، در 105 طی یک دوره 12-16 ساله، پس از آن اسید آمینه DNS جدا شد، با فلورسانس در تبادلات UV شناسایی شد.
قویترین روش حذف پروتئینهای DNS تحت هیدرولیز در حضور 5، 7 n است. HCl، در 105 طی یک دوره 12-16 ساله، پس از آن اسید آمینه DNS جدا شد، با فلورسانس در تبادلات UV شناسایی شد.


 روش S. Akabori l l هنگامی که پپتید (یا پروتئین) با هیدرازین بی آب در دمای 100-120 درجه سانتیگراد گرم می شود، پیوندهای پپتیدی با اسیدهای آمینه هیدرازیدها هیدرولیز می شوند، اسید آمینه C ترمینال از نظر ظاهری از بین می رود. از مخلوط دیده می شود و مشخص می شود:
روش S. Akabori l l هنگامی که پپتید (یا پروتئین) با هیدرازین بی آب در دمای 100-120 درجه سانتیگراد گرم می شود، پیوندهای پپتیدی با اسیدهای آمینه هیدرازیدها هیدرولیز می شوند، اسید آمینه C ترمینال از نظر ظاهری از بین می رود. از مخلوط دیده می شود و مشخص می شود:

 روش P. Edman l روش تجزیه پلی پپتیدی لانست با کمک فنیل ایزوتیوسیونات (FITC) توسط شیمیدان سوئدی P. V. Edman در سال 1950-1956 شکسته شد. روش ادمن امکان حذف مداوم باقی ماندههای اسید آمینه N ترمینال از فرم فنیل تیوهیدانتوئینها (PTH) را فراهم میکند. چرخه تخریب پوست شامل 3 مرحله است.
روش P. Edman l روش تجزیه پلی پپتیدی لانست با کمک فنیل ایزوتیوسیونات (FITC) توسط شیمیدان سوئدی P. V. Edman در سال 1950-1956 شکسته شد. روش ادمن امکان حذف مداوم باقی ماندههای اسید آمینه N ترمینال از فرم فنیل تیوهیدانتوئینها (PTH) را فراهم میکند. چرخه تخریب پوست شامل 3 مرحله است.
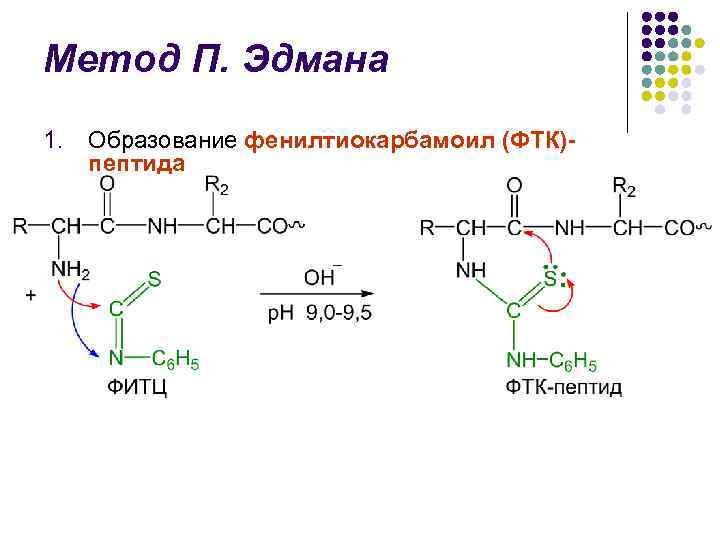



 روش P. Edman l l l شناسایی جداسازی PTG در ابتدا در روش ادمن است که طی سه ساعت گذشته کروماتوگرافی روی کاغذ انجام شد و سپس کروماتوگرافی کروی ریز بر روی سیلیکاژل و پلی آمید، کروماتوگرافی اصلی گاز طبیعی دستاوردهای بزرگ در زمینه ساختاری بود. پژوهش. ترتیب سنج ادمن و جی بیگ (از دنباله انگلیسی - sequence) - دستگاهی که با کارایی بالا، به طور خودکار باقیمانده اسید آمینه N ترمینال را به روش ادمن جدا می کند.
روش P. Edman l l l شناسایی جداسازی PTG در ابتدا در روش ادمن است که طی سه ساعت گذشته کروماتوگرافی روی کاغذ انجام شد و سپس کروماتوگرافی کروی ریز بر روی سیلیکاژل و پلی آمید، کروماتوگرافی اصلی گاز طبیعی دستاوردهای بزرگ در زمینه ساختاری بود. پژوهش. ترتیب سنج ادمن و جی بیگ (از دنباله انگلیسی - sequence) - دستگاهی که با کارایی بالا، به طور خودکار باقیمانده اسید آمینه N ترمینال را به روش ادمن جدا می کند.
 l برای تعیین ساختار پپتیدها و پروتئین ها، می توان موارد زیر را متراکم کرد: l L آنزیم هایی که کاتالیزور حذف باقی مانده های اسید آمینه N و C ترمینال هستند - آمینو کربوکسی پپتیدازها، روش های فیزیکوشیمیایی، طیف سنجی، طیف سنجی جرمی yu
l برای تعیین ساختار پپتیدها و پروتئین ها، می توان موارد زیر را متراکم کرد: l L آنزیم هایی که کاتالیزور حذف باقی مانده های اسید آمینه N و C ترمینال هستند - آمینو کربوکسی پپتیدازها، روش های فیزیکوشیمیایی، طیف سنجی، طیف سنجی جرمی yu
 تجزیه و تحلیل داده ها در مورد ساختار اولیه امکان توسعه اصول اساسی بیشتر را فراهم می کند. مشارکت احتمالی و تعداد کمی از پیوندهای دی سولفیدی در لنس پلی پپتیدی، ترکیبات مختلفی از اسیدهای آمینه ممکن است شناسایی شود.در پلی پپتیدها، همه دی پپتیدهای ممکن آشکار می شوند.
تجزیه و تحلیل داده ها در مورد ساختار اولیه امکان توسعه اصول اساسی بیشتر را فراهم می کند. مشارکت احتمالی و تعداد کمی از پیوندهای دی سولفیدی در لنس پلی پپتیدی، ترکیبات مختلفی از اسیدهای آمینه ممکن است شناسایی شود.در پلی پپتیدها، همه دی پپتیدهای ممکن آشکار می شوند.
 تجزیه و تحلیل داده ها در مورد ساختار اولیه به ما اجازه می دهد تا اصول اساسی زیر را توسعه دهیم: l L پوست یک پروتئین همگن فردی با ساختار اولیه منحصر به فرد مشخص می شود. اغلب، جایگزینی اسیدهای آمینه نه تنها منجر به تغییرات ساختاری می شود، بلکه منجر به تغییر در خواص فیزیکی و شیمیایی و عملکردهای بیولوژیکی می شود. این اصل شباهت ساختاری برای آنزیم های پروتئولیتیک کم - تریپسین، کیموتریپسین و غیره معمول است.
تجزیه و تحلیل داده ها در مورد ساختار اولیه به ما اجازه می دهد تا اصول اساسی زیر را توسعه دهیم: l L پوست یک پروتئین همگن فردی با ساختار اولیه منحصر به فرد مشخص می شود. اغلب، جایگزینی اسیدهای آمینه نه تنها منجر به تغییرات ساختاری می شود، بلکه منجر به تغییر در خواص فیزیکی و شیمیایی و عملکردهای بیولوژیکی می شود. این اصل شباهت ساختاری برای آنزیم های پروتئولیتیک کم - تریپسین، کیموتریپسین و غیره معمول است.

 در یک همجوشی غیر قطبی، انرژی رباط آب -CO НN- نزدیک به 16.7 کیلوژول بر مول می شود و تغییر قطبیت جریان میانی این انرژی را کاهش می دهد.
در یک همجوشی غیر قطبی، انرژی رباط آب -CO НN- نزدیک به 16.7 کیلوژول بر مول می شود و تغییر قطبیت جریان میانی این انرژی را کاهش می دهد.
 فعل و انفعالات متقابل هیدروفوبیک ماهیت آنتروپیک دارند زیرا مدافعان غیر قطبی وارد آب می شوند و سعی می کنند تماس خود را با آب محدود کنند مثلاً آب ساختار ساختاری خود را تجدید نمی کند و گویی یک گروه پریموس است واسطه هایی در آب وجود دارد. خوشه ای که حاوی حداقل انرژی است. مازاد
فعل و انفعالات متقابل هیدروفوبیک ماهیت آنتروپیک دارند زیرا مدافعان غیر قطبی وارد آب می شوند و سعی می کنند تماس خود را با آب محدود کنند مثلاً آب ساختار ساختاری خود را تجدید نمی کند و گویی یک گروه پریموس است واسطه هایی در آب وجود دارد. خوشه ای که حاوی حداقل انرژی است. مازاد

 فعل و انفعالات واندروالس شامل نیروهای پراکندگی، گرانش اتم ها و نیروهای انطباق متقابل پوسته های الکترونیکی آنها می شود. سهم انرژی در تماس با پوست کم است.
فعل و انفعالات واندروالس شامل نیروهای پراکندگی، گرانش اتم ها و نیروهای انطباق متقابل پوسته های الکترونیکی آنها می شود. سهم انرژی در تماس با پوست کم است.
 برهمکنش های یونی یا الکترواستاتیکی l برهمکنش های گروه های باردار l l برهمکنش های گروه های یون زایی که پیوندهای نمکی ایجاد می کنند انرژی پیوندهای نمکی می تواند به سطح آبگریز 41.9 k.J/mol برسد، اما مقدار آنها بسیار کم است. برهمکنش های یون-دوقطبی و دوقطبی-دوقطبی مشابه
برهمکنش های یونی یا الکترواستاتیکی l برهمکنش های گروه های باردار l l برهمکنش های گروه های یون زایی که پیوندهای نمکی ایجاد می کنند انرژی پیوندهای نمکی می تواند به سطح آبگریز 41.9 k.J/mol برسد، اما مقدار آنها بسیار کم است. برهمکنش های یون-دوقطبی و دوقطبی-دوقطبی مشابه
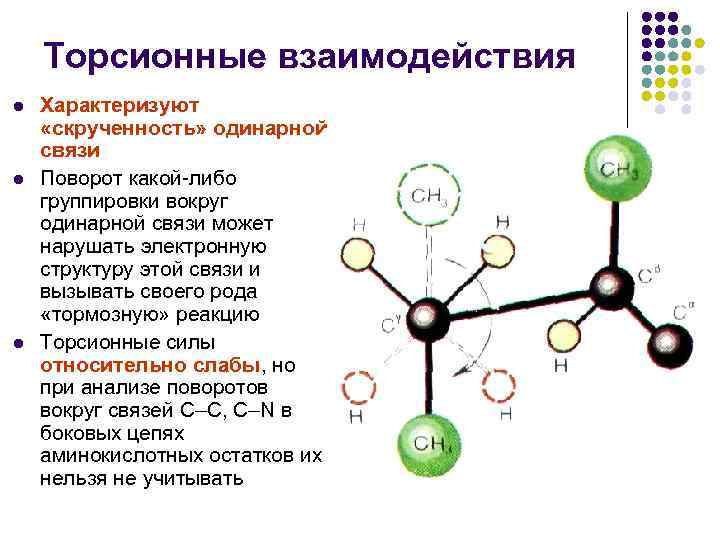 فعل و انفعالات پیچشی l l l مشخصه "پیچش" یک پیوند است. چرخش هر گروه بندی به دور یک پیوند می تواند ساختار الکترونیکی پیوند آن را از بین ببرد و نوعی واکنش "هلم" را ایجاد کند. واکنش پیچشی و نیروها به وضوح ضعیف هستند، اما زمانی که تجزیه و تحلیل چرخش در اطراف رباط їС، їN از لانست های زائد اسید آمینه اضافی را نمی توان اجتناب کرد.
فعل و انفعالات پیچشی l l l مشخصه "پیچش" یک پیوند است. چرخش هر گروه بندی به دور یک پیوند می تواند ساختار الکترونیکی پیوند آن را از بین ببرد و نوعی واکنش "هلم" را ایجاد کند. واکنش پیچشی و نیروها به وضوح ضعیف هستند، اما زمانی که تجزیه و تحلیل چرخش در اطراف رباط їС، їN از لانست های زائد اسید آمینه اضافی را نمی توان اجتناب کرد.

 ساختار ثانویه پروتئین... l l انبساط گسترده مقاطع گرد لانست پلی پپتیدی بدون نوع و ترکیب رادیکال های اسید آمینه. این توسط برهمکنش پیوندهای آبی بین گروه های پپتیدی هر دو لنست و گروه های مختلف ایجاد می شود. لانتسیوگیو
ساختار ثانویه پروتئین... l l انبساط گسترده مقاطع گرد لانست پلی پپتیدی بدون نوع و ترکیب رادیکال های اسید آمینه. این توسط برهمکنش پیوندهای آبی بین گروه های پپتیدی هر دو لنست و گروه های مختلف ایجاد می شود. لانتسیوگیو
 ساختار ثانویه یک پروتئین l l l هر بخش از یک مولکول پروتئین دارای ساختار ثانویه است.بعضی ساختار ثانویه عناصر تناوبی تر را در نظر می گیرند: - مارپیچ و - ساختار. اما در پروتئین ها بخش هایی که ساختار دارند در یک ساختار دقیق تر می شوند. به روشی ملایم، اگرچه ساختار جادار آن هر از گاهی مکرر و موتیف منظم از کسی انتقام نمی گیرد. قبل از آنها، مفهوم ساختار ثانویه را کاملاً درک می کنیم. 2 نوع ساختار ثانویه وجود دارد: منظم و نامنظم. مفهوم ساختار ثانویه برای تمام مولکول های پروتئین به طور کلی صدق نمی کند، بلکه برای چند بخش بزرگتر از کف استفاده می شود. لانست ایپپتیدی
ساختار ثانویه یک پروتئین l l l هر بخش از یک مولکول پروتئین دارای ساختار ثانویه است.بعضی ساختار ثانویه عناصر تناوبی تر را در نظر می گیرند: - مارپیچ و - ساختار. اما در پروتئین ها بخش هایی که ساختار دارند در یک ساختار دقیق تر می شوند. به روشی ملایم، اگرچه ساختار جادار آن هر از گاهی مکرر و موتیف منظم از کسی انتقام نمی گیرد. قبل از آنها، مفهوم ساختار ثانویه را کاملاً درک می کنیم. 2 نوع ساختار ثانویه وجود دارد: منظم و نامنظم. مفهوم ساختار ثانویه برای تمام مولکول های پروتئین به طور کلی صدق نمی کند، بلکه برای چند بخش بزرگتر از کف استفاده می شود. لانست ایپپتیدی
 ساختار ثانویه پروتئین l فعل و انفعالاتی که مهمترین نقش را در ساختار ثانویه تشکیل شده ایفا می کنند با موارد زیر مشخص می شوند: l l ویژگی های پیوند پپتیدی و برهمکنش های فضایی (- و - برش ها) نقش اصلی توسط پیوندهای آبی بین گروه های پپتیدی، که به صورت دوره ای در Lancus تکرار می شوند
ساختار ثانویه پروتئین l فعل و انفعالاتی که مهمترین نقش را در ساختار ثانویه تشکیل شده ایفا می کنند با موارد زیر مشخص می شوند: l l ویژگی های پیوند پپتیدی و برهمکنش های فضایی (- و - برش ها) نقش اصلی توسط پیوندهای آبی بین گروه های پپتیدی، که به صورت دوره ای در Lancus تکرار می شوند
 تثبیت ساختار ثانویه برای تشکیل اتصال دهنده های آب l تزریق پروتئین اضافی با آب آب می تواند به طور موثر برای تشکیل اتصال دهنده های آب رقابت کند: تزریق آب هنگام قالب گیری یک ساختار پروتئینی فشرده و جادار کاهش می یابد، رشد و به جای پیوندهای پپتیدی، افزایش سازگاری فعل و انفعالات آنها بنابراین، پایداری ساختار ثانویه در ساختار سوم فشرده
تثبیت ساختار ثانویه برای تشکیل اتصال دهنده های آب l تزریق پروتئین اضافی با آب آب می تواند به طور موثر برای تشکیل اتصال دهنده های آب رقابت کند: تزریق آب هنگام قالب گیری یک ساختار پروتئینی فشرده و جادار کاهش می یابد، رشد و به جای پیوندهای پپتیدی، افزایش سازگاری فعل و انفعالات آنها بنابراین، پایداری ساختار ثانویه در ساختار سوم فشرده

 -Spiral l در صخره های دهه 50 قرن XX. L. Pauling و R. Corey، بر اساس دادههای مربوط به ساختار کریستالهای اسیدهای آمینه و پپتیدهای ساده، به ترکیبهای دورهای احتمالی پلیپپتیدی لانژوگ پرداختند و به این نتیجه رسیدند که همگنترین ساختار توسط آنها -spi rally نامگذاری شده است.
-Spiral l در صخره های دهه 50 قرن XX. L. Pauling و R. Corey، بر اساس دادههای مربوط به ساختار کریستالهای اسیدهای آمینه و پپتیدهای ساده، به ترکیبهای دورهای احتمالی پلیپپتیدی لانژوگ پرداختند و به این نتیجه رسیدند که همگنترین ساختار توسط آنها -spi rally نامگذاری شده است.
 -Spiral انتخاب مکان بر اساس معیارهای زیر است: 1. ایجاد یک ساختار فشرده فشرده بدون خالی شدن و انتقاد بیش از حد از اتم ها 2. حداکثر اشباع ساختار با پیوندهای آبی از این ماده شیمیایی، بنابراین هندسه آنها نزدیک به خطی 3 بود. Dotrimannya بین افزایش دهنده های اتمی و گوشه ها و ببخشید پپتید L
-Spiral انتخاب مکان بر اساس معیارهای زیر است: 1. ایجاد یک ساختار فشرده فشرده بدون خالی شدن و انتقاد بیش از حد از اتم ها 2. حداکثر اشباع ساختار با پیوندهای آبی از این ماده شیمیایی، بنابراین هندسه آنها نزدیک به خطی 3 بود. Dotrimannya بین افزایش دهنده های اتمی و گوشه ها و ببخشید پپتید L
 مارپیچ l در ذهن این افراد، هم می تواند یک مارپیچ راست و هم یک مارپیچ چپ وجود داشته باشد، اما برای سمت راست، مارپیچ از نظر انرژی برجسته تر از چپ به نظر می رسد، زیرا لانس پپتیدی با اسیدهای آمینه L ایجاد می شود.
مارپیچ l در ذهن این افراد، هم می تواند یک مارپیچ راست و هم یک مارپیچ چپ وجود داشته باشد، اما برای سمت راست، مارپیچ از نظر انرژی برجسته تر از چپ به نظر می رسد، زیرا لانس پپتیدی با اسیدهای آمینه L ایجاد می شود.
 -Spiral l l ساختار مارپیچ زمانی ایجاد می شود که تمام اضلاع لانست پلی پپتیدی به دور رباط های ساده (i) بچرخد و دارای یک مقدار و علامت (نزدیک به 60) باشد، که منجر به پیچش گام به گام لانست می شود. رادیکال ها بیش از حد غیر اسیدی هستند که در حاشیه مارپیچ استوانه محلول ظاهر می شوند و بسته به ماهیت خود می توانند از ماهیت آبگریز یا آبدوست سطح استوانه ای اطمینان حاصل کنند.
-Spiral l l ساختار مارپیچ زمانی ایجاد می شود که تمام اضلاع لانست پلی پپتیدی به دور رباط های ساده (i) بچرخد و دارای یک مقدار و علامت (نزدیک به 60) باشد، که منجر به پیچش گام به گام لانست می شود. رادیکال ها بیش از حد غیر اسیدی هستند که در حاشیه مارپیچ استوانه محلول ظاهر می شوند و بسته به ماهیت خود می توانند از ماهیت آبگریز یا آبدوست سطح استوانه ای اطمینان حاصل کنند.
 - پارامترهای هندسی مارپیچ: l l l شعاع r = 2.3 Å (0.23 نانومتر) ارتفاع مارپیچ (تغییر مکان) در هر 1 اضافی d = 1.5 Å لبه مارپیچ (دوره هویت) P = 5.4 Å 1 چرخش مارپیچ 3، 6 اسید آمینه اضافی، همه - پیوندهای C=O مستقیم به سمت جلو، به انتهای C، و گروه های -N-H مستقیماً عقب هستند. در مارپیچ، گروه پوستی -NH توسط یک پیوند آبی به هم متصل می شود. به گروه -CO از اسید آمینه چهارم بیش از حد. znyogo (5 1 تماس)
- پارامترهای هندسی مارپیچ: l l l شعاع r = 2.3 Å (0.23 نانومتر) ارتفاع مارپیچ (تغییر مکان) در هر 1 اضافی d = 1.5 Å لبه مارپیچ (دوره هویت) P = 5.4 Å 1 چرخش مارپیچ 3، 6 اسید آمینه اضافی، همه - پیوندهای C=O مستقیم به سمت جلو، به انتهای C، و گروه های -N-H مستقیماً عقب هستند. در مارپیچ، گروه پوستی -NH توسط یک پیوند آبی به هم متصل می شود. به گروه -CO از اسید آمینه چهارم بیش از حد. znyogo (5 1 تماس)
 -Spiral l l تعداد بخشهای مارپیچی در پروتئین های کروی به طور قابل ملاحظه ای کم است (5 - 15 اسید آمینه اضافی، 3 - 4 چرخش مارپیچ)، در پروتئین های فیبریلی - بسیار گسترده است. لطفاً با مکان هایی که مازاد روشن است تماس بگیرید. تا سیستم ها قطع شوند. در این مرحله کل مارپیچ به مدت 20-30 فروکش می کند
-Spiral l l تعداد بخشهای مارپیچی در پروتئین های کروی به طور قابل ملاحظه ای کم است (5 - 15 اسید آمینه اضافی، 3 - 4 چرخش مارپیچ)، در پروتئین های فیبریلی - بسیار گسترده است. لطفاً با مکان هایی که مازاد روشن است تماس بگیرید. تا سیستم ها قطع شوند. در این مرحله کل مارپیچ به مدت 20-30 فروکش می کند

 -ساختار l l پیشنهاد شده توسط W. T. Astbury در سال 1941. بر اساس مطالعات ساختاری اشعه ایکس - کراتین پس از 10 سال، L. Pauling و R. Corey دریافتند که ساختار یا "ورق تاشو" به دلیل تثبیت رباطهای آب بین لنسینار مرتبط با بافتنی و زیگزاگی است. رباط های پپتیدی
-ساختار l l پیشنهاد شده توسط W. T. Astbury در سال 1941. بر اساس مطالعات ساختاری اشعه ایکس - کراتین پس از 10 سال، L. Pauling و R. Corey دریافتند که ساختار یا "ورق تاشو" به دلیل تثبیت رباطهای آب بین لنسینار مرتبط با بافتنی و زیگزاگی است. رباط های پپتیدی




 -ساختار l l تعداد اسیدهای آمینه اضافی در قسمتی از نیزه پپتیدی که ساختار - را ایجاد می کند از 3 تا 8 متغیر است. 6 عدد نیزه، گاهی اوقات تا 10 عدد
-ساختار l l تعداد اسیدهای آمینه اضافی در قسمتی از نیزه پپتیدی که ساختار - را ایجاد می کند از 3 تا 8 متغیر است. 6 عدد نیزه، گاهی اوقات تا 10 عدد
 -ساختار l l l گروه های اسیدی اسیدهای آمینه اضافی، زمانی که ساختار ایجاد می شود، توسط اضلاع مختلف سطح آنها آشکار می شود. سطح خود شکلی چین خورده دارد و چین ها توسط اتم های کربن مشخص می شوند. گروه های آلی، که از آنها حاصل می شود، شانه ایجاد می شود این به شما امکان می دهد سطوح طولانی پر از همان نوع رادیکال های بیولوژیکی (به عنوان مثال، آبگریز) سطح آبگریز توپ تا شده را قالب گیری و گسترش دهید، در تعامل با یکدیگر یا با برآمدگی های آبگریز مارپیچ ها. از هسته های آبگریز مولکولی داخلی، که به راحتی ساختار پروتئین را تثبیت می کند، شرکت کنید.
-ساختار l l l گروه های اسیدی اسیدهای آمینه اضافی، زمانی که ساختار ایجاد می شود، توسط اضلاع مختلف سطح آنها آشکار می شود. سطح خود شکلی چین خورده دارد و چین ها توسط اتم های کربن مشخص می شوند. گروه های آلی، که از آنها حاصل می شود، شانه ایجاد می شود این به شما امکان می دهد سطوح طولانی پر از همان نوع رادیکال های بیولوژیکی (به عنوان مثال، آبگریز) سطح آبگریز توپ تا شده را قالب گیری و گسترش دهید، در تعامل با یکدیگر یا با برآمدگی های آبگریز مارپیچ ها. از هسته های آبگریز مولکولی داخلی، که به راحتی ساختار پروتئین را تثبیت می کند، شرکت کنید.

 -Vigin l l هر دو مارپیچ و ساختار در پروتئین های کروی در بخش های کوتاه ارائه می شوند، به این معنی که بخشی از ساختار ثانویه پروتئین بر روی انواع مختلف حلقه ها قرار می گیرد، که به شما امکان می دهد جهت لانژوگ پپتید را تغییر دهید. عنصر ساختاری که به پلی پپتید اجازه می دهد 180 درجه بچرخد. 3 گروه پپتیدی، - -ویجین تثبیت شده توسط یک پیوند آبی تقریباً همیشه در سطح گلبول پروتئین ظاهر می شود، که اغلب نقش مهمی در برهمکنش آن با مولکول های دیگر دارد.
-Vigin l l هر دو مارپیچ و ساختار در پروتئین های کروی در بخش های کوتاه ارائه می شوند، به این معنی که بخشی از ساختار ثانویه پروتئین بر روی انواع مختلف حلقه ها قرار می گیرد، که به شما امکان می دهد جهت لانژوگ پپتید را تغییر دهید. عنصر ساختاری که به پلی پپتید اجازه می دهد 180 درجه بچرخد. 3 گروه پپتیدی، - -ویجین تثبیت شده توسط یک پیوند آبی تقریباً همیشه در سطح گلبول پروتئین ظاهر می شود، که اغلب نقش مهمی در برهمکنش آن با مولکول های دیگر دارد.


 l نتایج تجزیه و تحلیل ساختاری اشعه ایکس نشان داد که نحوه تا شدن مولکول پروتئین توسط توالی اسید آمینه آن تعیین می شود - gly, pro, asn Yakshcho با 6 اسید آمینه اضافی گروه بندی شده 4 برای ترکیب ایجاد مارپیچ، سپس این گروه اگر 3 مورد اضافی از 5 مورد گروه بندی شده با هم ترکیب شوند تا ساختار را ایجاد کنند، آنگاه این گروه بندی دانه ای است برای
l نتایج تجزیه و تحلیل ساختاری اشعه ایکس نشان داد که نحوه تا شدن مولکول پروتئین توسط توالی اسید آمینه آن تعیین می شود - gly, pro, asn Yakshcho با 6 اسید آمینه اضافی گروه بندی شده 4 برای ترکیب ایجاد مارپیچ، سپس این گروه اگر 3 مورد اضافی از 5 مورد گروه بندی شده با هم ترکیب شوند تا ساختار را ایجاد کنند، آنگاه این گروه بندی دانه ای است برای

 ساختار ثانویه (فوق ثانویه) پروتئین ها این سطح از سازماندهی مولکول های پروتئین با حضور مجموعه هایی از ساختارهای ثانویه در تعامل با یکدیگر مرتبط است.
ساختار ثانویه (فوق ثانویه) پروتئین ها این سطح از سازماندهی مولکول های پروتئین با حضور مجموعه هایی از ساختارهای ثانویه در تعامل با یکدیگر مرتبط است.

 مارپیچی فوق مارپیچی l l با چرخاندن مارپیچ های برگردان در پروتوفیبریل (کراتین) تشکیل می شود که به میکروفیبریل تبدیل می شود - مارپیچ ها توسط نیروهای واندروالس به هم فشرده می شوند.
مارپیچی فوق مارپیچی l l با چرخاندن مارپیچ های برگردان در پروتوفیبریل (کراتین) تشکیل می شود که به میکروفیبریل تبدیل می شود - مارپیچ ها توسط نیروهای واندروالس به هم فشرده می شوند.

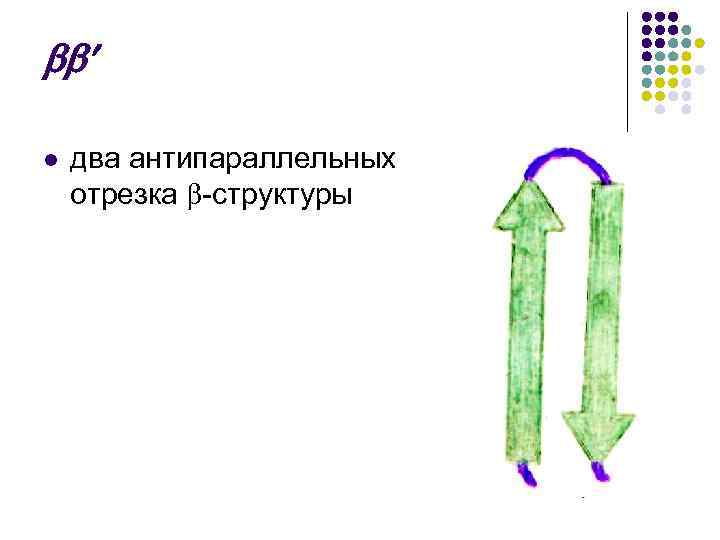
 x l بند، که از دو توپ موازی با مفاصل بین آنها تشکیل شده است، شبیه به: l توپ نامرتب - با
x l بند، که از دو توپ موازی با مفاصل بین آنها تشکیل شده است، شبیه به: l توپ نامرتب - با
 x l بند، که از دو توپ موازی با مفصل بندی بین آنها تشکیل شده است، به نظر می رسد: l l - مارپیچ - لنتس گذاشته شده طبق Rossman. دو قطعه متوالی متصل ساختار "زیور آلات یونانی" را ایجاد می کنند -
x l بند، که از دو توپ موازی با مفصل بندی بین آنها تشکیل شده است، به نظر می رسد: l l - مارپیچ - لنتس گذاشته شده طبق Rossman. دو قطعه متوالی متصل ساختار "زیور آلات یونانی" را ایجاد می کنند -
 بند x l که از دو توپ موازی با اتصالات بین آنها تشکیل شده است، به نظر می رسد: l - ساختار - - - زیگزاگ، "کلید گردو"
بند x l که از دو توپ موازی با اتصالات بین آنها تشکیل شده است، به نظر می رسد: l - ساختار - - - زیگزاگ، "کلید گردو"

 دامنه ها و یک به یک بخش های کوتاه لنس پلی پپتیدی که به آنها بخش های لولا می گویند دامنه های عملکردی می توانند از یک یا چند حوزه ساختاری تشکیل شوند.در تعدادی از آنزیم ها مرکز فعال بین دامنه ها جدا می شود.
دامنه ها و یک به یک بخش های کوتاه لنس پلی پپتیدی که به آنها بخش های لولا می گویند دامنه های عملکردی می توانند از یک یا چند حوزه ساختاری تشکیل شوند.در تعدادی از آنزیم ها مرکز فعال بین دامنه ها جدا می شود.



 3. + -پروتئین ها بخش هایی هستند که عمدتاً از مارپیچ و بخش هایی از توپ های ضد موازی انسولین ساخته شده اند.
3. + -پروتئین ها بخش هایی هستند که عمدتاً از مارپیچ و بخش هایی از توپ های ضد موازی انسولین ساخته شده اند.

 4. / -پروتئین ها l l - مارپیچ ها و ساختارها در طول مسیر لانکوگ تشکیل می شوند. بیشتر ساختارها (موازی) در قسمت مرکزی مولکول قرار دارند، جایی که این ساختارها در ظاهر پروانه خم می شوند (" ساختار پیچشی)، ایجاد یک "پایه" سفت و سخت، که سایر بخش های مولکول به آن متصل می شوند.
4. / -پروتئین ها l l - مارپیچ ها و ساختارها در طول مسیر لانکوگ تشکیل می شوند. بیشتر ساختارها (موازی) در قسمت مرکزی مولکول قرار دارند، جایی که این ساختارها در ظاهر پروانه خم می شوند (" ساختار پیچشی)، ایجاد یک "پایه" سفت و سخت، که سایر بخش های مولکول به آن متصل می شوند.


 دامنه ها l l l اندازه متوسط یک دامنه 100 تا 150 اضافی است که گویچه هایی با قطر تقریباً 2.5 نانومتر را نشان می دهد. ه- تشکیل ساختار سوم که در مراحل پایانی تشکیل گلبول اتفاق می افتد
دامنه ها l l l اندازه متوسط یک دامنه 100 تا 150 اضافی است که گویچه هایی با قطر تقریباً 2.5 نانومتر را نشان می دهد. ه- تشکیل ساختار سوم که در مراحل پایانی تشکیل گلبول اتفاق می افتد

 ساختار سوم پروتئین توزیع گسترده ای از مقاطع منظم و آمورف را در لانست پلی پپتیدی به عنوان یک کل مشخص می کند که به دلیل برهمکنش رادیکال های بیولوژیکی است و در نوع و ترکیب آنها نهفته است. بنابراین، ساختار سوم توصیف می کند. آرایش فضایی همه مولکول های پروتئین، زیرا آنها توسط یک ماده پلی پپتیدی که مستقیماً با شکل مولکول های پروتئین مرتبط است، ساخته شده اند، که می تواند از رشته ای تا نخ مانند متفاوت باشد.
ساختار سوم پروتئین توزیع گسترده ای از مقاطع منظم و آمورف را در لانست پلی پپتیدی به عنوان یک کل مشخص می کند که به دلیل برهمکنش رادیکال های بیولوژیکی است و در نوع و ترکیب آنها نهفته است. بنابراین، ساختار سوم توصیف می کند. آرایش فضایی همه مولکول های پروتئین، زیرا آنها توسط یک ماده پلی پپتیدی که مستقیماً با شکل مولکول های پروتئین مرتبط است، ساخته شده اند، که می تواند از رشته ای تا نخ مانند متفاوت باشد.
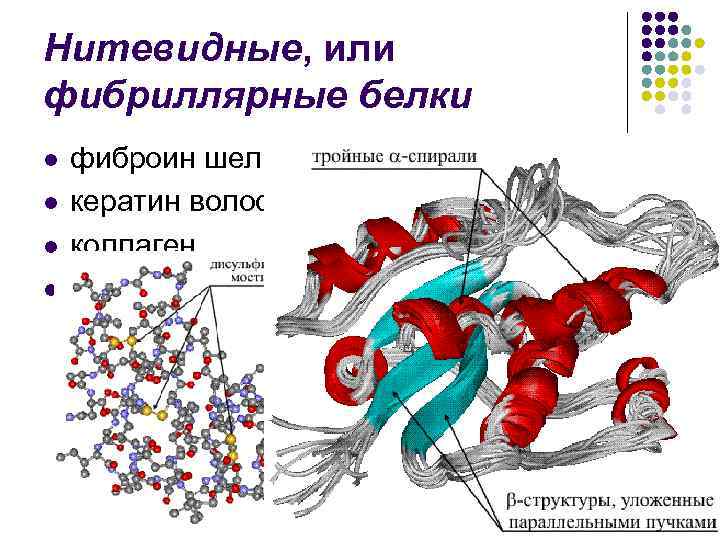 پروتئین های نخ مانند یا فیبریلار l l فیبروین بخیه کراتین مو، شاخ، تجمع کلاژن و غیره.
پروتئین های نخ مانند یا فیبریلار l l فیبروین بخیه کراتین مو، شاخ، تجمع کلاژن و غیره.

 ساختار سوم پروتئین اساس عملکرد پروتئین است که نیاز به سازماندهی فضایی دقیق مجموعه های بزرگ ایجاد شده در غیاب اسیدهای آمینه اضافی دارد. پروتئین (دناتوره شدن) باید تا زمانی که توانایی عملکرد خود را از دست بدهد انجام شود
ساختار سوم پروتئین اساس عملکرد پروتئین است که نیاز به سازماندهی فضایی دقیق مجموعه های بزرگ ایجاد شده در غیاب اسیدهای آمینه اضافی دارد. پروتئین (دناتوره شدن) باید تا زمانی که توانایی عملکرد خود را از دست بدهد انجام شود
 ساختار سوم پروتئین l l l ثبات ساختار سوم در سیستم برهمکنش های غیرکووالانسی در وسط گلبول پروتئین نهفته است (؟) برخی از پروتئین ها علاوه بر این توسط پیوندهای کووالانسی - دی سولفیدی - Glanzug = RTln تثبیت می شوند. K = Nchain – T S lanceug Nchain 0 (S
ساختار سوم پروتئین l l l ثبات ساختار سوم در سیستم برهمکنش های غیرکووالانسی در وسط گلبول پروتئین نهفته است (؟) برخی از پروتئین ها علاوه بر این توسط پیوندهای کووالانسی - دی سولفیدی - Glanzug = RTln تثبیت می شوند. K = Nchain – T S lanceug Nchain 0 (S
 ساختار سوم پروتئین l Spivvіdnosti Chain i -T Schip در موارد زیر است: l تعداد پیوندهای غیر کووالانسی که هنگام ورود مولکول های پروتئین به گلبول نصب می شوند و بعد از لنسر پلی پپتیدی به دما بستگی دارد. زی زروستانیام تی |ت س| زود یا دیر حرکت | N| که ساختار باز ثبات را از دست می دهد - دناتوره شدن حرارتی پروتئین
ساختار سوم پروتئین l Spivvіdnosti Chain i -T Schip در موارد زیر است: l تعداد پیوندهای غیر کووالانسی که هنگام ورود مولکول های پروتئین به گلبول نصب می شوند و بعد از لنسر پلی پپتیدی به دما بستگی دارد. زی زروستانیام تی |ت س| زود یا دیر حرکت | N| که ساختار باز ثبات را از دست می دهد - دناتوره شدن حرارتی پروتئین
 ساختار سوم پروتئین l l هنگامی که گلبول بلعیده می شود، آنتروپی لانس پپتید تغییر می کند، اما همزمان افزایش آنتروپی مبدأ - آب وجود دارد که این نقش کلیدی در تثبیت ساختار سوم دارد.
ساختار سوم پروتئین l l هنگامی که گلبول بلعیده می شود، آنتروپی لانس پپتید تغییر می کند، اما همزمان افزایش آنتروپی مبدأ - آب وجود دارد که این نقش کلیدی در تثبیت ساختار سوم دارد.
 ساختار سوم پروتئین l l آب یک عامل ساختاری بی اهمیت و درجه بالا است (؟) گروه های قطبی پروتئین به راحتی در ساختارهای یخ مانند آب ادغام می شوند و پیوندهای آبی از آن وارد می شوند. فضاهای خالی خود را که توسط مولکول ها ایجاد می شود را اشغال می کنند.
ساختار سوم پروتئین l l آب یک عامل ساختاری بی اهمیت و درجه بالا است (؟) گروه های قطبی پروتئین به راحتی در ساختارهای یخ مانند آب ادغام می شوند و پیوندهای آبی از آن وارد می شوند. فضاهای خالی خود را که توسط مولکول ها ایجاد می شود را اشغال می کنند.
 ساختار سوم پروتئین l l اگر قطعات گروه های غیر قطبی پروتئین نتوانند از مولکول "کره" خارج شوند، یک گلبول تشکیل می شود که در آن بخش قابل توجهی (نه کمتر از ½) از گروه های آبگریز در تماس با آنها ظاهر می شود. اب این مستلزم نصب کنتاکت های آبگریز، کنتاکت های واندروالس است. نیروها در آنتروپی پروتئین کاهش می یابد به دنبال تشکیل گلبول، بخش قابل توجهی از ویسکوزیته آب و برهمکنش های آبگریز گروه های پروتئینی با مولکول های آب از بین می رود. هرج و مرج باقی مانده افزایش می یابد و آنتروپی آب افزایش می یابد
ساختار سوم پروتئین l l اگر قطعات گروه های غیر قطبی پروتئین نتوانند از مولکول "کره" خارج شوند، یک گلبول تشکیل می شود که در آن بخش قابل توجهی (نه کمتر از ½) از گروه های آبگریز در تماس با آنها ظاهر می شود. اب این مستلزم نصب کنتاکت های آبگریز، کنتاکت های واندروالس است. نیروها در آنتروپی پروتئین کاهش می یابد به دنبال تشکیل گلبول، بخش قابل توجهی از ویسکوزیته آب و برهمکنش های آبگریز گروه های پروتئینی با مولکول های آب از بین می رود. هرج و مرج باقی مانده افزایش می یابد و آنتروپی آب افزایش می یابد

 ساختار چهارتایی یک پروتئین... l l l ساختار چهارتایی در پروتئین های الیگومری یافت می شود که مولکول های آن از دو یا چند لنج پلی پپتیدی تشکیل شده است که به صورت غیر کووالانسی متصل شده اند. مجموعه ای از گلبول ها) به عنوان یک مولکول واحد عمل می کند، در عین حال پوست گلبول های ترکیب شده استقلال قابل توجهی را حفظ می کند.
ساختار چهارتایی یک پروتئین... l l l ساختار چهارتایی در پروتئین های الیگومری یافت می شود که مولکول های آن از دو یا چند لنج پلی پپتیدی تشکیل شده است که به صورت غیر کووالانسی متصل شده اند. مجموعه ای از گلبول ها) به عنوان یک مولکول واحد عمل می کند، در عین حال پوست گلبول های ترکیب شده استقلال قابل توجهی را حفظ می کند.

 ساختار چهارتایی یک پروتئین l l زیرواحدهایی که ساختار چهارتایی یک پروتئین را ایجاد می کنند می توانند کاملاً با عملکرد واقعی آنها متفاوت باشند - پروتئین های هترومری که به شما امکان می دهد تعدادی از عملکردهای متقابل وابسته به یکدیگر را در یک ساختار ترکیب کنید و یک مولکول غیرعملکردی کف l l App ایجاد کنید. پروتئین کیناز: زیرواحد C مسئول فعالیت آنزیمی است و زیرواحد R تنظیم کننده است.در پروتئین های همومریک، زیرواحدها یکسان هستند.
ساختار چهارتایی یک پروتئین l l زیرواحدهایی که ساختار چهارتایی یک پروتئین را ایجاد می کنند می توانند کاملاً با عملکرد واقعی آنها متفاوت باشند - پروتئین های هترومری که به شما امکان می دهد تعدادی از عملکردهای متقابل وابسته به یکدیگر را در یک ساختار ترکیب کنید و یک مولکول غیرعملکردی کف l l App ایجاد کنید. پروتئین کیناز: زیرواحد C مسئول فعالیت آنزیمی است و زیرواحد R تنظیم کننده است.در پروتئین های همومریک، زیرواحدها یکسان هستند.
 ساختار چهارتایی پروتئین l تماس های بین زیر واحدی – سیستم برهمکنش های غیرکووالانسی قبلاً توسعه یافته است.ساختار چهارتایی کمتر و درجه سوم است، به این معنی که تماس های آبگریز کمتری وجود دارد.
ساختار چهارتایی پروتئین l تماس های بین زیر واحدی – سیستم برهمکنش های غیرکووالانسی قبلاً توسعه یافته است.ساختار چهارتایی کمتر و درجه سوم است، به این معنی که تماس های آبگریز کمتری وجود دارد.

 توابع یک ساختار یک چهارم 1. ترکیب بسیاری از عملکردهای مرتبط به هم در یک ساختار واحد 2. عملکرد معماری 1. شکل دادن ساختارهای جادار به شکل تاشوهای یکنواخت برای اطمینان از ظرفیت خاص و عملکردی پروتئین (فریتین) 2. خلاصه ای از واکنش های آنزیمی بعدی 3. تشکیل عملکردی
توابع یک ساختار یک چهارم 1. ترکیب بسیاری از عملکردهای مرتبط به هم در یک ساختار واحد 2. عملکرد معماری 1. شکل دادن ساختارهای جادار به شکل تاشوهای یکنواخت برای اطمینان از ظرفیت خاص و عملکردی پروتئین (فریتین) 2. خلاصه ای از واکنش های آنزیمی بعدی 3. تشکیل عملکردی
 توابع ساختار چهارم 3. اطمینان از برهمکنش های متعدد پروتئین با ساختارهای توسعه یافته پروتئین های اتصال به DNA - دیمرها (؟) 4. عملکرد تنظیمی. انتقال اثر (آسیب به ساختار سوم در اثر تعامل با بستر) از یک زیر واحد به زیر واحد دیگر منتقل می شود که منجر به بازسازی تمام ساختارهای چهارتایی می شود.
توابع ساختار چهارم 3. اطمینان از برهمکنش های متعدد پروتئین با ساختارهای توسعه یافته پروتئین های اتصال به DNA - دیمرها (؟) 4. عملکرد تنظیمی. انتقال اثر (آسیب به ساختار سوم در اثر تعامل با بستر) از یک زیر واحد به زیر واحد دیگر منتقل می شود که منجر به بازسازی تمام ساختارهای چهارتایی می شود.


 1. پروتئین پوست فردی با ساختار منحصربهفردی مشخص میشود که منحصر به فرد بودن عملکرد آن را تضمین میکند.
1. پروتئین پوست فردی با ساختار منحصربهفردی مشخص میشود که منحصر به فرد بودن عملکرد آن را تضمین میکند.
 ادبیات Berezov T. T.، Korovkin B. F. شیمی بیولوژیکی. - M.: پزشکی، 1983 Metzler D. Biochemistry. واکنش های شیمیایی در سلول های زنده در 3 جلد - T. volume 1, 2. - M.: Svit, 1980 Ovchinnikov Yu. A. Bioorganic chemistry. - M.: Prosvitnitstvo، 1987 مبانی بیوشیمی / ویرایش. A. A. Anisimova. - M.: Vishcha Shkola، 1986 Rhys E.، Strenberg M. از سلول ها به اتم ها. معرفی مصور زیست شناسی مولکولی. - M.: Svit، 1988 Stepanov V. M. زیست شناسی مولکولی. ساختار و عملکرد پروتئین ها - M.: مدرسه Vishcha، 1996 Pilipovich Yu. B. مبانی بیوشیمی. - م.: مدرسه ویشچا، 1993
ادبیات Berezov T. T.، Korovkin B. F. شیمی بیولوژیکی. - M.: پزشکی، 1983 Metzler D. Biochemistry. واکنش های شیمیایی در سلول های زنده در 3 جلد - T. volume 1, 2. - M.: Svit, 1980 Ovchinnikov Yu. A. Bioorganic chemistry. - M.: Prosvitnitstvo، 1987 مبانی بیوشیمی / ویرایش. A. A. Anisimova. - M.: Vishcha Shkola، 1986 Rhys E.، Strenberg M. از سلول ها به اتم ها. معرفی مصور زیست شناسی مولکولی. - M.: Svit، 1988 Stepanov V. M. زیست شناسی مولکولی. ساختار و عملکرد پروتئین ها - M.: مدرسه Vishcha، 1996 Pilipovich Yu. B. مبانی بیوشیمی. - م.: مدرسه ویشچا، 1993


